Basement Membrane Disruption by Tumor Necrosis Factor-α in the μSiM-hNVU
Note: a postscript regarding an assay to determine the activity of our MMP-2/MMP-9 inhibitor was added to this post on June 14, 2021.
Introduction
The blood-brain barrier (BBB) plays an important role in maintaining brain chemistry by restricting the permeability of small molecules and cells. Inflammatory diseases may disrupt the BBB and promote the development of neurological disorders [1]. One potential mechanism of BBB disruption is the degradation of the extracellular matrix proteins that make up the endothelial basement membrane (BM). The BM is composed primarily of collagen IV and laminin networks that are linked by nidogen, perlecan, and heparan sulfate proteoglycans [2]. These proteins provide a support structure for endothelial cells, and it is believed that cellular adhesion to collagen IV is part of a pathway responsible for upregulation of the tight junction protein claudin-5 [3]. The tight junctions are the key to limiting endothelial permeability, since they restrict paracellular migration of small molecules. Thus, the BM helps maintain the barrier function of the BBB in two ways: its networks act as physical impediments to molecular diffusion, and they help sustain tight junctions. BM proteins are degraded by matrix metalloproteinase-9 (MMP-9, also known as Type IV Collagenase or Gelatinase B), which is upregulated by pro-inflammatory signals such as tumor necrosis factor-α (TNF-α) [4]. This could be an important link between inflammation and BBB disruption.
MMP-9 Treatment Times
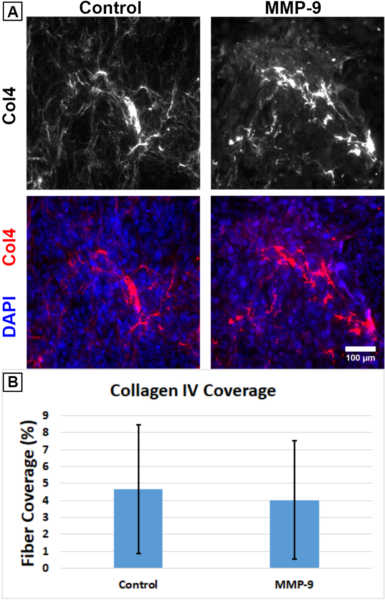
Previously, we treated co-cultures of human Cerebral Microvascular Endothelial Cells (hCMEC/D3; Sigma-Aldrich cat. no. SCC066) and primary Human Brain Vascular Pericytes (HBVPs; ScienCell cat. no. 1200) on nanoporous μSiM membranes with 0.1 μg/mL active human recombinant MMP-9 (Sigma-Aldrich cat. no. PF140) over a ~6.25 hr period and reported a reduction in collagen IV fiber coverage. We expressed a lack of confidence in these earlier results due to the small number of surviving experimental replicates (several of the μSiM membranes broke during fixation and staining), so we repeated this study with a larger number of devices.
HBVPs were seeded on the underside of the μSiM’s nanoporous membrane followed by hCMEC/D3s in the top well 2 days later. Experiments were conducted 4 days after hCMEC/D3 seeding. 0.1 μg/mL MMP-9 was added to half of the devices, and the rest received PBS mixed in media as a control. Following 6.25 hr, the devices were fixed and stained for collagen IV (Fig. 1A). To determine the percentage of area covered by collagen IV fibers, the immunofluorescence images were automatically contrast-adjusted and binarized using Fiji (Available: https://imagej.net/Fiji). Unlike the prior experiment, there was no indication of a difference between the two conditions (Fig. 1B). It is probable that MMP-9 requires more time to have a noticeable impact on collagen IV degradation.
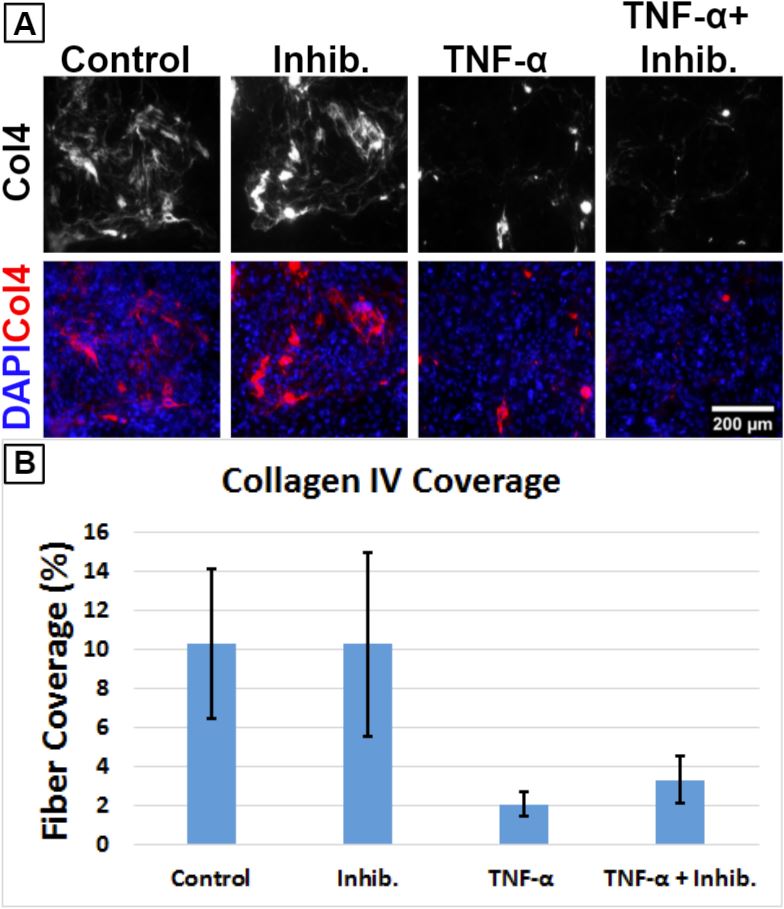
TNF-α Treatment
While we were setting up the MMP-9 experiment discussed above, we were simultaneously piloting studies with the proinflammatory cytokine TNF-α. After hCMEC/D3 seeding, the μSiMs were cultured for 6 days. The cells were treated with 20 ng/mL TNF-α (Sigma-Aldrich, cat. no. H8916) with or without the inclusion of 1 μg/mL MMP-2/MMP-9 inhibitor (Sigma-Aldrich, cat. no. 444241) for ~6.25 hr. Since the pathway leading to MMP-9 upregulation is believed to be TNF-α’s primary method of influencing the BM, we introduced the inhibitor to see if this would be able to attenuate any effects caused by TNF-α. However, 6.25 hr was insufficient to produce any noticeable effect on collagen IV coverage (Fig. 2).

Increasing the treatment time to 24 hr produced remarkably more apparent effects on collagen IV (Fig. 3). Fiber coverage dropped from around 10% down to 2% following exposure to TNF-α. Treatment with the MMP inhibitor alone did not affect collagen IV fiber coverage. The MMP inhibitor failed to completely protect against the effects of TNF-α. It is possible that a higher concentration of inhibitor is required or that the cells should be primed by the inhibitor in advance of TNF-α introduction. Alternatively, TNF-α could be upregulating collagen-digesting products other than MMPs.

Discussion
While the results from the previous MMP-9 experiment were not reproducible, the promising outcome from the 24 hr TNF-α treatment demonstrated that collagen IV degradation can still be monitored effectively in the μSiM device. However, the TNF-α experiments suffer from the same limitation as our earlier work with MMP-9: there are few replicates for each condition. We intend to repeat the 24 hr TNF-α treatment using a larger number of devices. It may also be useful to prime the cells with the MMP inhibitor for different periods of time prior to TNF-α treatment to see how this impacts the effects of proinflammatory signaling. Furthermore, we can extend the TNF-α treatment period up to 4 days to see if collagen IV is able to begin its recovery. These longer experiments may give us the opportunity to explore the model’s permeability to FITC-dextran over time, and it would be interesting to see how this correlates with collagen IV coverage.
Postscript: MMP Inhibitor Activity
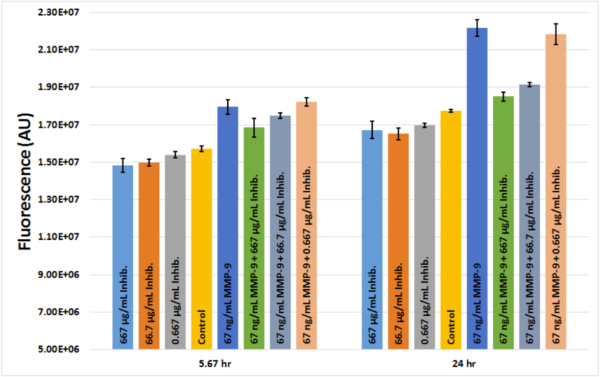
Our MMP-2/MMP-9 inhibitor did not obviously demonstrate protection against TNF-α in the experiment described above. It was unclear whether the inhibitor was active or if its concentration was simply too low. In order to determine if the inhibitor was working properly, we utilized dye-quenched gelatin conjugated to fluorescein (Invitrogen cat. no. D12054) gifted to us by the McGrath Lab which was previously used to confirm the activity of our MMP-9. When an active gelatinase or collagenase digests the gelatin, the quenching agent is removed. This results in increased fluorescence that can be measured with a plate reader. The protocol is available here. 67 ng/mL MMP-9 was added to the gelatin and was treated with three different concentrations of the inhibitor (667 μg/mL, 66.7 μg/mL, and 0.667 μg/mL). Fluorescence was measured at 5.67 hr and 24 hr. The inhibitor worked in a dose-dependent manner, though the lowest concentration was unable to attenuate the effects of the MMP-9 (Fig. 4). Since the amount of active MMP-9 secreted by hCMEC/D3s during TNF-α stimulation is currently unknown, it is not clear what concentration of the inhibitor will be needed in future experiments.
References
[1] X. Cong and W. Kong, “Endothelial tight junctions and their regulatory signaling pathways in vascular homeostasis and disease,” Cellular Signalling, vol. 66, p. 109485, 2020/02/01/ 2020.
[2] C. Leclech, C. F. Natale, and A. I. Barakat, “The basement membrane as a structured surface – role in vascular health and disease,” Journal of Cell Science, vol. 133, p. jcs239889, 2020.
[3] T. Osada, et al., “Interendothelial claudin-5 expression depends on cerebral endothelial cell-matrix adhesion by β(1)-integrins,” Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism, vol. 31, pp. 1972-1985, 2011/10// 2011.
[4] G. A. Rosenberg, E. Y. Estrada, J. E. Dencoff, and W. G. Stetler-Stevenson, “Tumor necrosis factor-α-induced gelatinase B causes delayed opening of the blood-brain barrier: an expanded therapeutic window,” Brain Research, vol. 703, pp. 151-155, 1995/12/12/ 1995.
Very interesting and a lot of experiments here. Can you edit the post to let us know what the sample sizes were?
Does the literature suggest the 1 μg/mL concentration for the inhibitor? I would be tempted to try a higher dose before concluding that TNF is working through a different pathway. We should also try MMP9 ELISA.
The post has been updated with sample sizes.
I could not find a clear answer regarding appropriate inhibitor concentrations. I agree that higher doses may be worth examining. An ELISA for MMP-9 would also be useful.
We confirmed that the MMP inhibitor is active in a dose-dependent manner, so perhaps larger concentrations will be needed in order to attenuate the effects of TNF-α. A postscript with more information has been added to the end of this post.
Do you know if the pericyte number under the membrane drops with the loss of collagen IV? This is thought to happen when the BBB breaks down and reproducing it here would be awesome validation of our model.