Fibronectin and Friends: Confirming the Presence of Multiple Vascular Basement Membrane Components in the μSiM-hNVU
Introduction
The blood-brain barrier (BBB) plays an important role in maintaining brain chemistry by restricting the permeability of small molecules and cells. Inflammatory diseases may disrupt the BBB and promote the development of neurological disorders [1]. One potential mechanism of BBB disruption is the degradation of the extracellular matrix proteins that make up the vascular basement membrane (BM). The BM is composed primarily of collagen IV and laminin networks that are linked by nidogen, perlecan, and heparan sulfate proteoglycans [2]. These proteins provide a support structure for endothelial cells, and it is believed that cellular adhesion to collagen IV is part of a pathway responsible for upregulation of the tight junction protein claudin-5 [3]. The tight junctions are the key to limiting endothelial permeability since they restrict paracellular diffusion of small molecules. Thus, the BM helps maintain the barrier function of the BBB in two ways: its networks act as physical impediments to molecular diffusion, and they help sustain tight junctions. BM proteins are degraded by matrix metalloproteinase-9 (MMP-9, also known as Type IV Collagenase or Gelatinase B), which is upregulated by pro-inflammatory signals such as tumor necrosis factor-α (TNF-α) [4]. This could be an important link between inflammation and BBB disruption. Our goal is to model this effect using the μSiM-hNVU.
We have accomplished a lot of prior work with collagen IV, including our recent posts here and here. This post summarizes our latest efforts to confirm the presence of fibronectin and laminin in our BBB model. In addition, we took a first look at BM deposition by iPSC-derived BBB cells and also revisited the effects of cell treatment with pro-inflammatory cytokines.
The majority of this work has been performed by co-culturing two cell types: a human cerebral microvascular endothelial cell line (hCMEC/D3; Sigma-Aldrich cat. no. SCC066) and primary human brain vascular pericytes (HBVPs; ScienCell cat. no. 1200). Co-culturing was accomplished by coating the trench side of the μSiM membrane with 2 μg/cm2 poly-L-lysine prior to “trench-down” device assembly, followed by inverted seeding of the HBVPs. hCMEC/D3s were seeded in the 25 µg/cm2 collagen I + 5 µg/cm2 fibronectin-coated top well 1-2 days later. Devices were subsequently maintained for ~7 days prior to starting experiments to ensure that there was sufficient time for BM protein deposition.
iPSC work was performed by Molly McCloskey at the University of Rochester. iPSCs were differentiated into Extended Endothelial Cell Culture Method-Brain Microvascular Endothelial Cell (EECM-BMEC)-like cells as well as isogenic brain pericyte-like cells (BPLCs). μSiM co-cultures were generated by seeding the BPLCs on the underside of the membrane followed a day later by seeding the EECM-BMEC-like cells on the collagen IV- and fibronectin-coated top well. These co-cultures were maintained for 7 days prior to fixation and staining.
Results
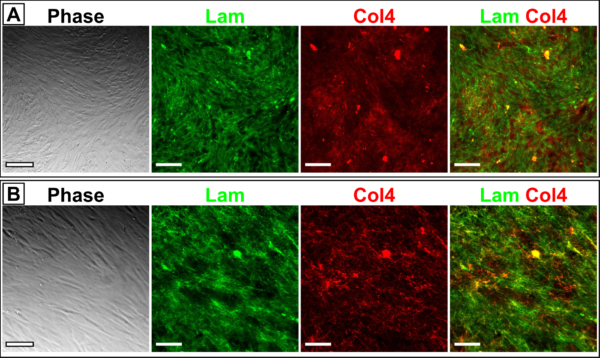
We performed tests of our fibronectin and laminin antibodies with HBVPs grown on tissue culture polystyrene prior to staining cells in the μSiMs. Fibronectin seemed to label extracellular fibers without issue, but there appeared to be lots of intracellular laminin (Fig. 1). One interesting takeaway from this initial experiment was that laminin seemed to be colocalized with collagen IV (Fig. 1B). It was clear that we would need to use an alternate staining approach in order to exclusively label extracellular laminin. We tried a couple different ways to reduce plasma membrane permeability including fixation in a formaldehyde + sucrose solution and omitting Tx-100, but in the end the most successful method was to live-stain the cells for laminin prior to fixation (Fig. 2). This did not seem to block too many cell attachment sites, since the HBVPs were still adherent and confluent following completion of the live-staining protocol.

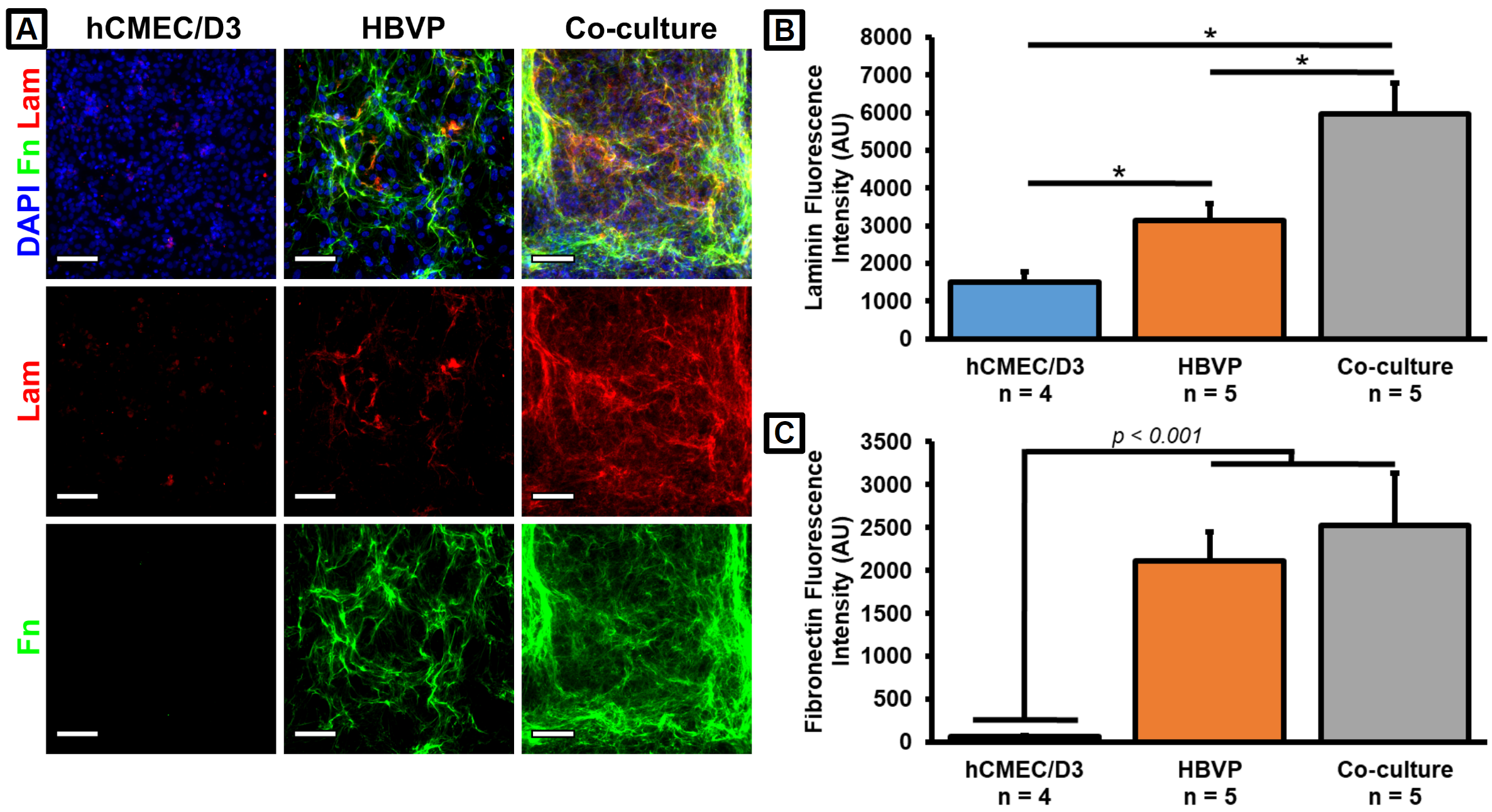
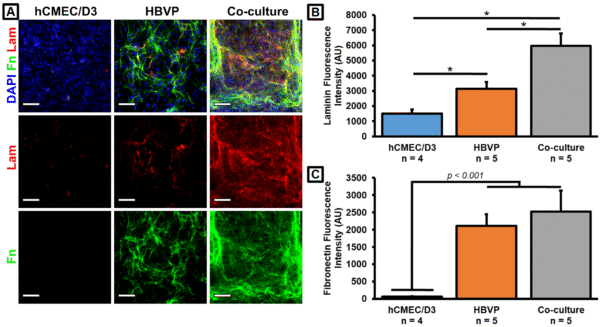
We had previously determined that HBVPs were responsible for the majority of collagen IV production in our μSiM co-cultures. We examined similar trends for both fibronectin and laminin (Fig. 3). There may be a small amount of laminin production by the hCMEC/D3s, and the cell types may act in either an additive or synergistic fashion such that the co-cultures produce more total laminin than the HBVP monocultures (Fig. 3B). However, it was clear that almost all fibronectin was being produced by the HBVPs (Fig. 3C). Note that 5 μg/cm2 fibronectin was used to coat the top well of the devices. However, staining a cell-free coated membrane revealed no substantial fluorescence signal (not shown), and we have concluded that all of the detected fibronectin was deposited by the HBVPs. These results confirmed that our cells were capable of producing two major BM constituents in addition to the previously observed collagen IV in our BBB model.
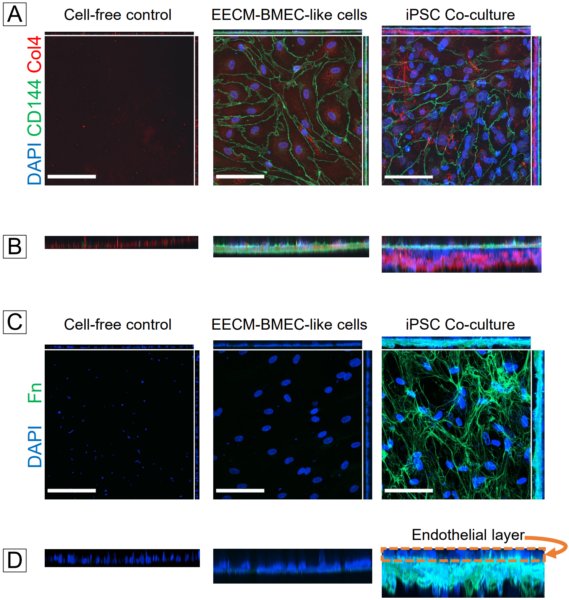
In our previous post, we used confocal microscopy to demonstrate that collagen IV was deposited on the abluminal side of the hCMEC/D3s in co-cultures, thus mimicking the physiological orientation of the BM. We also wanted to ensure that this was the case for our iPSC-derived EECM-BMEC-like cells and BPLCs. Immunofluorescent labeling and confocal microscopy did indeed show that the endothelial-specific VE-cadherin stain was concentrated in a layer above the collagen IV in our co-cultures (Fig. 4A,B). There was little collagen IV apparent in EECM-BMEC-like cell monocultures, indicating that the BPLCs were probably the main contributors of collagen IV. We also stained a cell-free coated control since the membranes were coated with collagen IV and fibronectin to support the EECM-BMEC-like cells. The coated control exhibited little collagen IV fluorescence compared to the co-culture devices. Similar results were obtained with fibronectin (Fig. 4C,D). While we were unable to stain for an endothelial-specific marker in the fibronectin images, the “naked nuclei” of the endothelial layer are plainly visible above the fibronectin layer in our co-culture image.
We also conducted a preliminary test to stain laminin in the iPSC-derived co-cultures. It appeared that there was indeed laminin present (Fig. 5), but we do not currently have any monoculture devices or cell-free coated membrane control devices to compare with.

Moving back to the hCMEC/D3 and HBVP co-cultures, we sought to demonstrate that our model could recapitulate the inflammatory response resulting in BM degradation. We had previously treated our devices with 20 ng/mL Tumor Necrosis Factor Alpha (TNF-α) with or without 100 µg/mL MMP-2/MMP-9 Inhibitor I for 24 hr on several occasions, but only had a few successes prior to 2022. This and other experimental results led us to conclude that our TNF-α may have lost activity over the 1.5 years spent in our -20°C freezer. Recently, we repeated the experiment with a fresh batch of TNF-α and stained for collagen IV and laminin (Fig. 6). While there was no significant difference between conditions, the trends for collagen IV matched our expectations with a 31.3% average drop in fluorescence intensity due to the TNF-α treatment that was rescued by co-treatment with the MMP inhibitor (Fig. 6B). The results for fibronectin were a little less clean due to an apparent drop in fluorescence intensity after treatment solely with the MMP inhibitor, but otherwise the trends for fibronectin were similar to collagen IV (Fig. 6C).
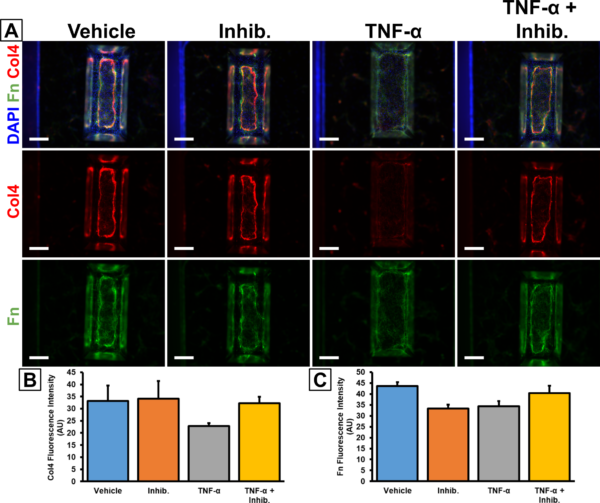
A. Immunofluorescence images were taken at 4x magnification on a Keyence BZ-X microscope. Scale bars = 500 μm. B. Barplot of collagen IV fluorescence intensity from the porous membrane region of the immunostained images. C. Barplot of fibronectin fluorescence intensity from the porous membrane region of the immunostained images. n = 3 (n = 4 for TNF-α treatment condition). Error bars = SEM.
In figure 6, it is apparent that the collagen IV and fibronectin deposition seemed to take on an elongated “oval shape” on the μSiM membrane (Fig. 6A). This phenomenon had been observed previously, as well as with some of the iPSC-derived co-cultures (Fig. 7). We hypothesize that BM deposition is restricted by the walls of the μSiM trench. Furthermore, it has been observed that the pericytes will sometimes contract away from the edges of the membrane after reaching confluency. This might mean that the BM is not continuous on the underside of the μSiM membrane, thus diminishing the physiological character of the BBB model. One possible solution may be to flip the chip such that the pericytes are grown on the flat side. Pericyte culture in multiwell plates generally results in continuous coverage by the deposited ECM proteins (Fig. 1,2), so it is likely that this will also occur on the flat side of the μSiM membranes. However, this would mean that the endothelial cells would need to be grown on the trench side and it is unknown if the endothelial interactions with the trench walls would disrupt monolayer and barrier formation.

Conclusions
We are able to stain for BM components including collagen IV, laminin, and fibronectin, though live-staining must be performed in order to avoid labeling intracellular laminin. It is unclear exactly which laminin isoforms are present in our BBB model since our antibody is polyclonal, but we could potentially determine this in the future if we want to ensure that we have laminin subtypes specifically associated with the brain microvasculature. Pericytes appear to be the main contributors of fibronectin and laminin in our model, which is similar to what had previously been observed with collagen IV. This seems to hold true for the iPSC-derived BPLCs in addition to the primary HBVPs. BM proteins are concentrated below the endothelial layer in our iPSC co-cultures, mimicking the physiological orientation of the BM around the vasculature. 24 hr treatment with TNF-α resulted in loss of collagen IV and fibronectin fluorescence, though this change was not significant. Different treatment lengths and doses of TNF-α should be evaluated to see if we can better recreate the in vivo response to inflammation. Finally, we are concerned that the μSiM trench walls are constricting the pericytes and limiting their deposition of BM proteins. In order to obtain a continuous layer of extracellular matrix proteins it may be necessary to culture the pericytes on the flat side of the membrane, though endothelial culture in the trench may negatively impact barrier formation.
References
[1] X. Cong and W. Kong, “Endothelial tight junctions and their regulatory signaling pathways in vascular homeostasis and disease,” Cellular Signalling, vol. 66, p. 109485, 2020/02/01/ 2020.
[2] C. Leclech, C. F. Natale, and A. I. Barakat, “The basement membrane as a structured surface – role in vascular health and disease,” Journal of Cell Science, vol. 133, p. jcs239889, 2020.
[3] T. Osada, et al., “Interendothelial claudin-5 expression depends on cerebral endothelial cell-matrix adhesion by β(1)-integrins,” Journal of cerebral blood flow and metabolism : official journal of the International Society of Cerebral Blood Flow and Metabolism, vol. 31, pp. 1972-1985, 2011/10// 2011.
[4] G. A. Rosenberg, E. Y. Estrada, J. E. Dencoff, and W. G. Stetler-Stevenson, “Tumor necrosis factor-α-induced gelatinase B causes delayed opening of the blood-brain barrier: an expanded therapeutic window,” Brain Research, vol. 703, pp. 151-155, 1995/12/12/ 1995.
Fantastic progress. Flipping the chip worth trying. I wonder if we are getting some preferential PC migration up and down the long axis of the membrane. Diana Hudecz has done flip-chip permeability studies for years because it improves imaging. Her data indicates there is no issue with forming proper barriers.