High Throughput Multiplexed Digital-Molecular-Counting Blood-Brain-Barrier on a Chip System (dμSIM plate)
- Background
Tissue-on-a-chip systems have been a promising means of mimicking and understanding the complex and convoluted functions of the in vivo blood-brain barrier (BBB). These semipermeable borders of endothelial cells are highly selective in order to protect the fragile central nervous system (CNS) from bloodborne pathogens or inflammatory signals and are known to have a huge impact on the phenotypes of resident CNS cells e.g., the microglia, astrocytes. The BBB also restricts the penetration of drugs which results in the low success rate in the development of CNS therapeutics. Alzheimer drugs (Aricept®, Exelon®,Razadyne®) did well with mouse models, but failed in later human clinical trials because of the unique impermeability of the human BBB structure. Induced pluripotent stem cells (iPSCs) have become a popular choice to be implemented in these tissue-on-a-chip systems to reconstruct a representative human BBB. However, the cell differentiation and even growth in BBB systems takes long periods of time and suffer from large batch to batch variations. These prolonged sample preparation time and low repeatability between batches of cells hinder the progress of the BBB research. Current tissue-on-a-chip platforms focus more on maximizing the resemblance of the system to in vivo biological environments which leads to complex microfluidic structures and ultimately results in a low-throughput setting. Additionally, in current publishings of BBB-on-a-chip research, only the trans-endothelial electrical resistance (TEER) sensors have been implemented as a characterization of the constructed BBB and there are currently no immunosensors being integrated. The lack of a sensitive, multiplexed, and high throughput BBB platform prohibits the progression of adoption of BBB-on-a-chip into applications like pathway studies and drug discovery. Herein, we present a high throughput multiplexed Digital-Molecular-Counting Blood-Brain-Barrier-on-a-Chip System namely the dμSIM for efficient parallel cytokine secretomic phenotyping of the BBB.
- Method
The duSIM plate (duSIM) method is similar to the our previously published digital Phenoplate (Small 2021, 17, 2101743) where compartmentalizable microwell array sensors are prepatterned in arrayed formats on a glass substrate and cell secreted information will be captured and stored on the chip for downstream high throughput assay labeling and digital molecular counting readout. (Figure 1) Instead of the microbead-based format utilized in the dPP, the duSIM uses a newly developed beadless method which prevents bead loss during more layer debonding, liquid removal, and mixing processes required in the duSIM assay. The beadless digital sensor also has advantages in increased linear dynamic range, increased analyte concentration resolution and also further limits cross contamination between markers (3 plex in this work) as antibodies are immobilized in the microwells directly. Furthermore, the glass duSIM chip that was used to characterize the brain side secretion can be striped with piranha cleaning and a bead-based digital assay can be performed on the same chip to measure the supernatant of the blood side of the coordinating BBB enabling using one wafer (chip) for up to 56×3plex digital measurements.
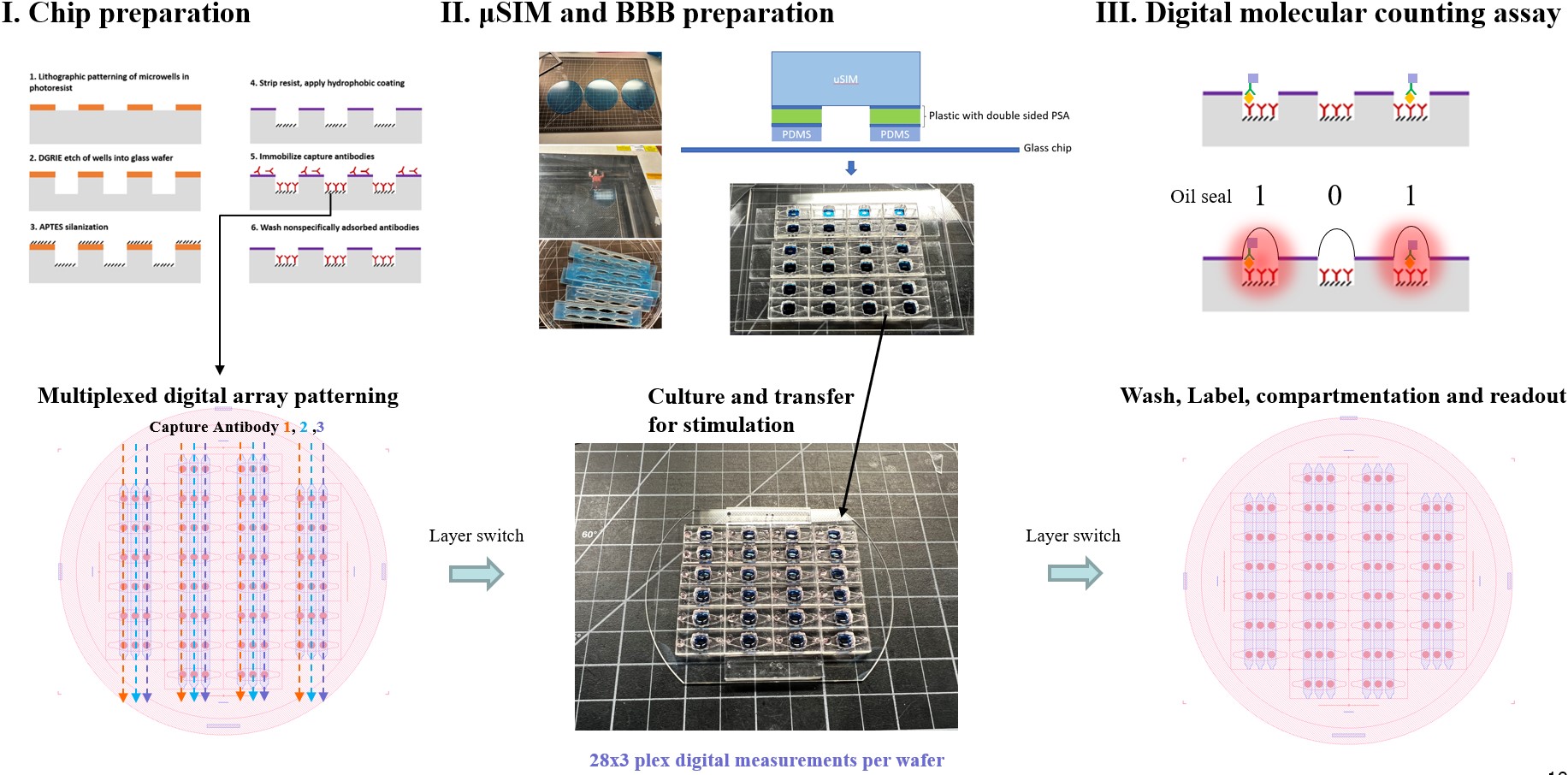
Figure 1. Schematic of the integrated digital counting uSIM process.
- Results and Discussions
COMSOL simulation of the concentration profile in our past post has proven that mixing is required for reproducibility and a back-and-forth oscillation mixing is sufficient to homogenize the bottom chamber. (Figure 2)

Figure 2. Oscillation mixing yields a homogenous cytokine distribution in the bottom channel
We have chosen MCP1, IL6 and TNFα as the three markers to simultaneously monitor in the brain side of the duSIM and calibration curves using both the bead-less and bead-based methods were successfully generated and plotted in Figure 3. The digital sensor linear dynamic range was optimized to be able to fit within the typical secretion range from the BBB tissues.

Figure 3. 3-plexed calibration curves using the bead-less (top three plots) and the bead-based (bottom plot) methods
We use primary mouse brain endothelial cells to reconstruct mouse BBB in this work to avoid the difficulties and instabilities in the differentiating endothelial cells from human stem cells. The brain endothelial cells typically reach confluency 3 days after plating and form tight junctions on day 4 or day 5. We then transfer the devices from a plain glass to our digital molecular counting chip for further stimulation and cytokine secretomic phenotyping. We have used various concentrations of LPS and cytokines and S100A8/A9 as the stimulant. Below are typical brightfield images of the duSIM imaging window and some tight junction marker (ZO1 and CLDN5) immunostaining results. (Figure 4.) We recognize that the cells in the fluorescent staining were not a monolayer which may have resulted in our difficulties in distinguishing between the LPS and non LPS stimulated groups. Additionally, we were not able to use ICAM1 as an activation marker even after switching antibodies from 3 vendors. We believe that mouse brain endothelial cells cultured in vitro might express really low levels of ICAM1 and we are therefore trying VCAM1 as an alternative activation imaging indicator to complement our cytokine profiling results. 
Figure 4. Representative brightfield and fluorescent staining images of the mouse brain endothelial cells cultured and stimulated in the duSIM
The 3-plex cytokine measurement results (Figure 5.) upon LPS stimulation exploited an interesting result as the BBB secret much higher than basal levels (3000pg/ml) of the chemokine MCP1 back to the blood but not to the brain side. On the contrary, for IL6 and TNFa, the BBB secret towards the brain side, but not to the blood side with TNFa secreting only ~20pg/ml when stimulated and 10pg/ml at basal. The IL6 secretion is also highly secreted towards the brain when being stimulated by LPS (5000~10000pg/ml) as compared to the blood (1000~2000pg/ml) and the basal (30~140pg/ml on both sides of the BBB)
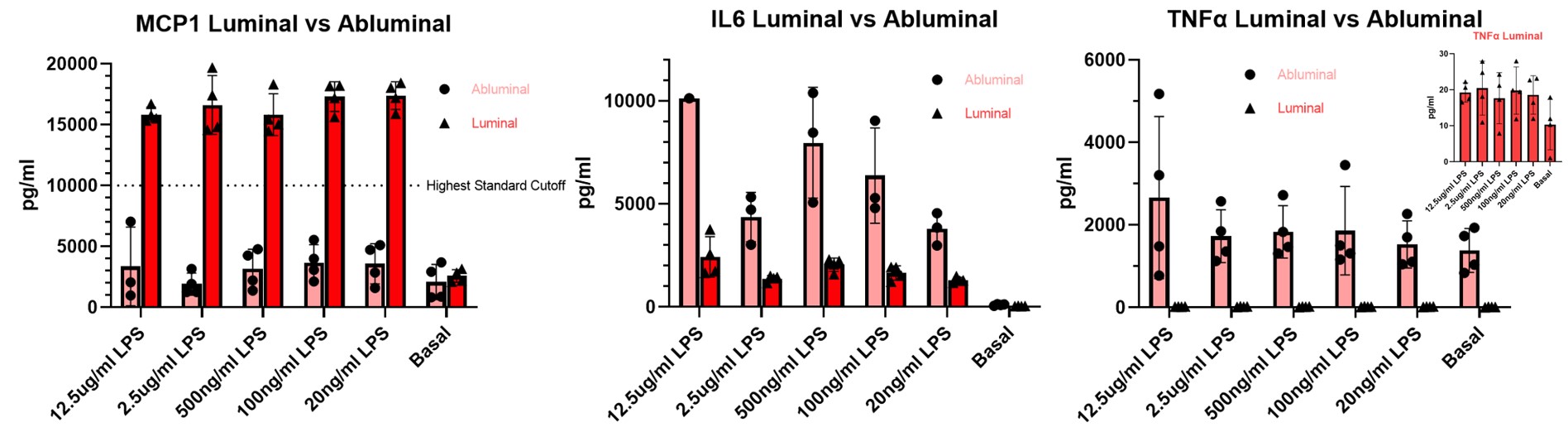
Figure 5. 3-plex cytokine (MCP1, IL6, TNFa) secretomic phenotyping upon various LPS concentrations stimulation
For the cytokine and S100A8/A9 stimulated experiments, we observed a high baseline of MCP1 secretion and gradual decrease of secretion on the brain side as stimulation concentration increased. The blood side concentration didn’t vary much across different dosages. IL6 concentrations were very low on both sides of the BBB which potentially indicates the under-stimulation at this level of dosage. Interestingly, we observed IP10 suppressing effect when co-stimulating 100pg/ml of cytokines with S100A8/A9. Extensive and repeated experiments will be needed to verify this phenomenon. When the concentrations of the blood and brain side are plotted on the same scale, we also observed interesting secretion polarization where the brain endothelial cells secret more inflammatory protein towards the brain when exposed to elevated cytokine levels from the blood.

Figure 6. 3-plex cytokine(MCP1, IL6, IP10) secretomic phenotyping upon various cytokine and S100A8/A9 concentrations stimulation
- Conclusion
The development of this platform will offer BBB researchers a tool to study multimodal (immunohistochemistry and 3-plexed cytokine secretion in this work) responses to various stimulants at scale. Other than some experimental attempts related to staining with the new VCAM1 antibodies and some SEM morphology characterization work of the duSIM for a publication, our next step is to try to measure cytokine stimulation responses from the brain endothelial cells in less than 2~4 hours of stimulation (currently 24hrs) to explore if we can pick up fast changes further utilizing our ultrasensitive capability of our digital sensors.