Human Tendon-on-Chip (hToC), a current research tool and future screening platform
The Burden of Tendon Injuries on HealthCare:
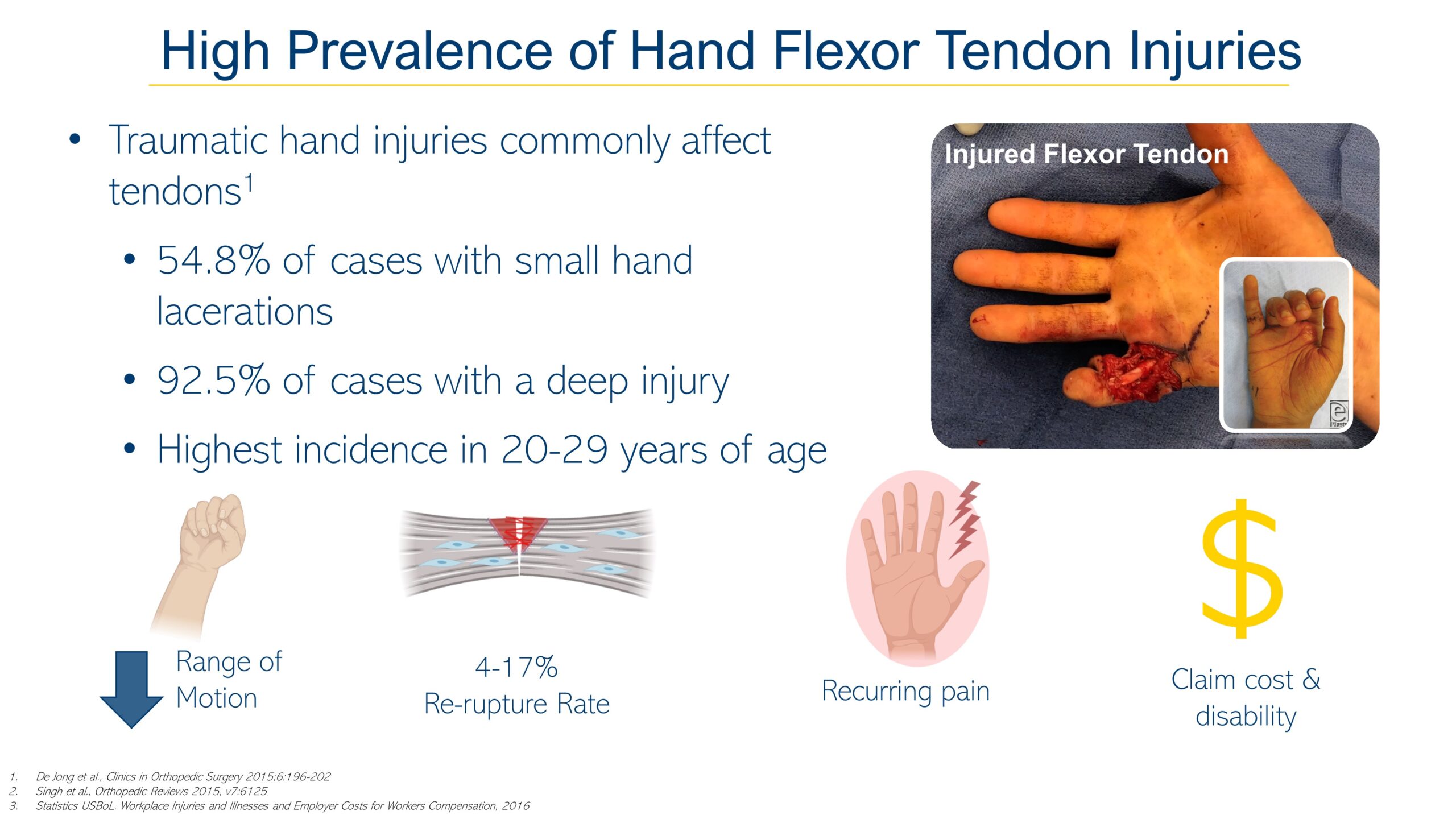
Acute and chronic tendon injuries commonly affecting the working-age population are frequently work-related and have negative socioeconomic outcomes such as productivity loss, income loss, and unemployment(O’Hara et al., 2020). Tendon injuries are often multifaceted and can be characterized by inflammation and degenerative changes in tendon structures collectively known as tendinopathies. Musculoskeletal conditions come in second to spinal pain and global volume of years lived with disability (Statistics, 2018). Both chronic and acute tendon injuries heal through the formation of a fibrovascular scar that never attains the gross, histological, or mechanical characteristics of native tendon(Galatz et al., 2015;Lin et al., 2018). The major injuries typically involve multiple tissues including the Achilles, Patellar, Quadriceps, Hamstring, Supraspinatus (rotator cuff), and hand and wrist flexor tendons. The socioeconomic consequences of tendon injuries can be evaluated by measuring outcomes in return to work and lost workdays as they translate into the value-based healthcare defined by the patient. According to the Bureau of Labor Statistics (BLS) the average hand injury claim can cost over  7.5K(Statistics, 2016). Despite the prevalence of tendon injuries, disease-modifying therapies have been hindered due to the incomplete understanding of fundamental tendon development, disease pathogenesis and healing. Developing targeted biological therapies that focus on regenerating tendon tissues to pre-injury condition will lower the economic burden associated with tendon injuries, which today are vastly addressed with non-biological, long-term treatments often administered over months/years.
7.5K(Statistics, 2016). Despite the prevalence of tendon injuries, disease-modifying therapies have been hindered due to the incomplete understanding of fundamental tendon development, disease pathogenesis and healing. Developing targeted biological therapies that focus on regenerating tendon tissues to pre-injury condition will lower the economic burden associated with tendon injuries, which today are vastly addressed with non-biological, long-term treatments often administered over months/years.
The frequent occurrence of tendon injuries has increased interest in interventional clinical trials. There are currently 126 active clinical trials on procedural drug or behavioral interventions for tendon disorders (Medicine). Biological and behavioral interventions in these studies include physical therapies, platelet-rich plasma biological treatments, chemical compounds, and innovative surgical procedures. Success metrics in these interventions are limited to compositional and structural changes of the tendon determined through assessments by the medical practitioner. Although these metrics provide activity guidelines for the patient, they do not provide information for the degree of healing in the tendon which can be established using identified serum biomarkers. We posit that the gap in knowledge of tendon healing pathobiology hinders the development of biologically relevant biomarkers and that novel human microphysiological systems (hMPS), or tissue-on-chip, can address this critical need by using clinical trials on-a-chip to de-risk human clinical trials. Current disease models of musculoskeletal conditions are restricted to animal models with limited translational value and simplified two-dimensional (2D) in vitro platforms that lack much of the physiology present in in vivo tendon(Kitta et al., 2018).
Here in a human tendon-on-a-chip (hToC), human derived endothelial cells line the lower channel compartment and a tenocyte seeded hydrogel rests on the top compartment separated by an ultrathin and highly permeable porous silicon membrane. Fibrosis is induced in the tendon hydrogel through dose dependent addition of TGF-β1, a crucial player in the induction and progression of fibrosis(Qiao et al., 2018). An increase in markers of cell senescence have been linked to fibrotic diseases where upregulation of SASPs and γH2AX are present as indicators(Kuo and Yang, 2008;Cárdenes et al., 2018). Myofibroblasts are the main cell types responsible for physiological reconstruction of connective tissues, such as tendon, after injury and in generating pathological tissue deformations that are characteristic of fibrosis(Hinz, 2007). Fibroblast are differentiated into activated myofibroblast to commence remodeling and most vastly identified through expression of alpha-smooth muscle actin (α-SMA)(Roach et al., 2014;Lu et al., 2018;Woodcock et al., 2019).
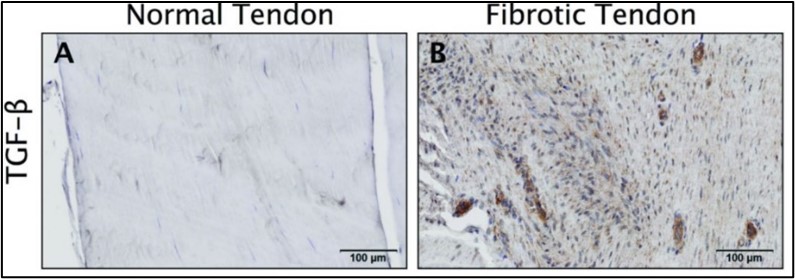
TGF-β1 has been established as a key regulator in the development of fibrotic diseases as it is highly involved in different signaling pathways and cross-talks between many signaling cascades(Eldred and Wormstone, 2015). In homeostatic conditions TGF-β1 restores native tissue architecture, yet in excess can compromise normal organ function(Branton and Kopp, 1999),(Li et al., 2004) as has been associated in fibrotic diseases of tendon(Farhat et al., 2012), lung(Schafer et al., 2017), heart(Li et al., 2004), and kidney(Qiao et al., 2018). Figure 2 depicts the upregulated fibrotic effects of TGF-β1 in fibrotic human tendon from research performed in our lab(Freeberg et al., 2018). The role of inflammation as a supporter or inducer of fibrosis and ultimately scar formation is not completely understood. Inflammation is thought to drive fibrosis through the activation of interleukins from either fibroblasts or macrophages (Borthwick, 2016). Specifically, interleukin-13 (IL-13) propagates tendon scarring via TGF-β1 released from activated macrophages. While recent advancements have demonstrated involvement of the inflammatory pathways in tendon healing, not much is understood about the pathway’s driving mechanisms(Thankam et al., 2019). Naïve macrophages (Mφs) classically activated (M1) Mφs, or alternatively activated (M2) Mφs represent the dynamic immune population that modulate the tendon regenerative and remodeling potential(Lin et al., 2018). Incorporation of early-stage pro-inflammatory innate immune cells, particularly Mφs, into the hToC elucidates the role of inflammatory mediators in inducing fibrosis in the hydrogel. Successful establishment of communication between Mφs, tendon fibroblasts and endothelial cells permits for novel understandings of how paracrine secretions induce the senescent phenotype in tendon fibroblast, a subject not yet investigated in the field.
Peritendinous Vascular Fibrosis Model:
Based on these observations and driven to develop a screening platform that identifies effective therapeutic targets for fibrovascular scar formation, we arrive at the proposed disease model. The main cell types involved are Endothelial Cells (ECs), Tenocytes (tendon-derived fibroblasts) and Macrophages all derived from human sources. We take advantage of collaborations with the University of Rochester Medical Center (URMC) and obtain human tendon tissue from which we isolate tendon fibroblasts and incorporate into the model.

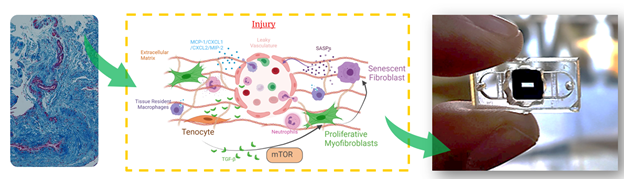
Figure 3: Current disease model of peritendinous vascular fibrosis in tendon. Left, H&E-stained histology section of human injured tendon, blood vessels stained in red and collagen matrix stained in blue. Right, proposed disease model based on human observations and physiological histology. In short, we propose that proliferative myofibroblast are induced by direct and paracrine interactions with resident and infiltrating macrophages, which ultimately lead to senescent fibroblasts and the release of SASPs. These perpetuate the feedback signaling involving TGF-beta and mTOR leading to chronic inflammation and fibrosis that manifests as adhesions. We believe that the elevation of specific SASP profiles can be used to assess biological targets that prevent or modulate peritendinous adhesions.
The proposed disease model, Figure 3, incorporates the current understanding of tendon fibrosis. Briefly, immune cells are recruited upon injury to the tendon tissue through infiltrating vasculature and initiate the inflammatory cascade. TGF-β released by macrophages in the tissue differentiate resident fibroblasts into activated myofibroblasts that secrete extracellular matrix proteins leading to the formation of a fibrous scar. These myofibroblasts polarize into a senescent phenotype as the fibrotic scar progresses in the tissue. The senescent myofibroblasts are no longer secreting matrix nor remodeling proteins but rather enter their role as signaling cells secreting Senescence-Associated Secreted Proteins (SASP). The constant secretion of SASP are thought to induce unresolved chronic inflammation leading to an un-remodeled tissue which has lost is pre-injury mechanical properties and is highly prone to re-injury. Nonetheless, much is still not understood in the healing response of tendon, such as the roles of circulating vs tissue-resident macrophages in the initiation and progression of fibrosis and alternative roles of senescent cells and SASP. Yet, investigating this cytokine milieu will give insight into effective therapeutic targets that can dampen the chronic inflammatory response and ultimately restore tendon function to patients’ post-injury.
RESULTS:
The following sections outline the steps we have taken thus far to design, develop and validate the peritendinous vascular fibrosis model.
The modular Human Tendon-on-Chip (hToC)
the hToC platform was built as extension of the µSiM platform (Figure 4). Specifically the lower compartment (Component 2) was re-engineered to provide more volume ~100 µL for the tendon hydrogel and engineered with contains two horizontally suspended bars which act as anchors to hold the hydrogel in place despite tenocyte driven contraction. The porous membrane contains 5um pores dispersed over a nanoporous background to allow for paracrine signaling and extravasation of larger migratory cells between the top well and bottom channel. As another device innovation, we take advantage of reversible nature of the pressure sensitive adhesive (PSA) used to join the two components to enable ‘one-sided’ or ‘joined’ investigations into the tissue model. Future studies will also take advantage of the 96-well format patterning of the for downstream high-throughput assays to power clinical trials-on-a-chip.

Modeling Peritendinous Vascular Fibrosis
To mimic peritendinous vascular fibrosis we must represent the interplay between vasculature and tendon tissue. We seed endothelial cells in the top chamber on the apical side of the membrane and place a primary tenocyte laden collagen hydrogel in the bottom channel, in contact with the basal side of the porous membrane (Figure 5). In panel 4C we see the tissues spatial distribution through a confocal z-stack of the endothelial cell monolayer and the primary human tenocytes suspended in the collagen hydrogel. This interaction between endothelial cells and tenocytes permits the investigation of immune components that infiltrate during fibrosis from the infiltrating vasculature. The platform is the first to incorporate a vascular component and cross talk of 3 cell types to model tendon scar. It is also compatible with many biological assays including individual access to each tissue for endpoint analysis, real-time visual access to each tissue and cytokine assays of the supernatant.

Figure 5: Left, cross-sectional view of hToC chambers including 2 fluidic access ports, a top well that exposes the porous membrane, a bottom channel that houses the tendon-tissue hydrogel and two static anchors horizontally suspended in bottom channel. A) Endothelial cell monolayer on membrane stained with junction protein VE-Cadherin (yellow). B) Tenocytes embedded in collagen hydrogel stained with F-Actin (green). C) Confocal z-stack of collagen hydrogel in bottom channel and ECs on membrane.
To first establish our model, we treated the system with 10 ng/mL Transforming Growth Factor-β1 (TGF- β1), attributed as the “Master regulator of fibrosis” in most fibrotic responses. This concentration was determined through the analysis of mouse serum 7-days post tendon injury. Experimentally, TGF-β1 serum-free media (XVIVO10, LONZA Cat#04-380Q) is introduced to both the top and bottom components. To isolate the responses of the individual cells and validate the effectiveness of our multi-culture system to model peritendinous adhesions various cellular combinations were tested and can be seen outlined in figure 6. The outputs we use to verify the induction of a fibrotic response in our model were aligned with in vivo, both in human and mouse, responses of fibrotic tissue and are outlined in the table below.
| In vivo symptom | hToC Measured Output | Expected Result |
| Presence of scar-formation depicted through irregular matrix formation and increased collagen secretion. |
1. Bulk-tissue contraction 2. α-Smooth Muscle Actin+ myofibroblasts |
1. Decreased area contraction as fibrosis begins 2. Increased α-SMA in fibrotic condition as fibroblasts differentiate into myofibroblasts |
| Unresolved chronic inflammation |
Senescence in Fibroblasts through 1. Nuclear staining of γH2AX 2. P16/P21 Expression 3. Secretory senescent profile (SASPs) |
1. Increased γH2AX in fibrotic state 2. Increased P16/P21 expression in fibrotic state 3. Pro-inflammatory cytokine elevated in inflammatory state |

The first combination tested was the tenocytes (tendon fibroblasts) in the bottom chamber with our Type I/III collagen hydrogel (R&D Systems, Cat# 5026). Since no ECs were required for this experiment, a top reservoir was utilized instead of the top component with the porous membrane, all volumes remained the same. Confirming our proposed disease model, we observed bulk tissue contraction indicating a pro-fibrotic response from the tenocytes (figure 7).
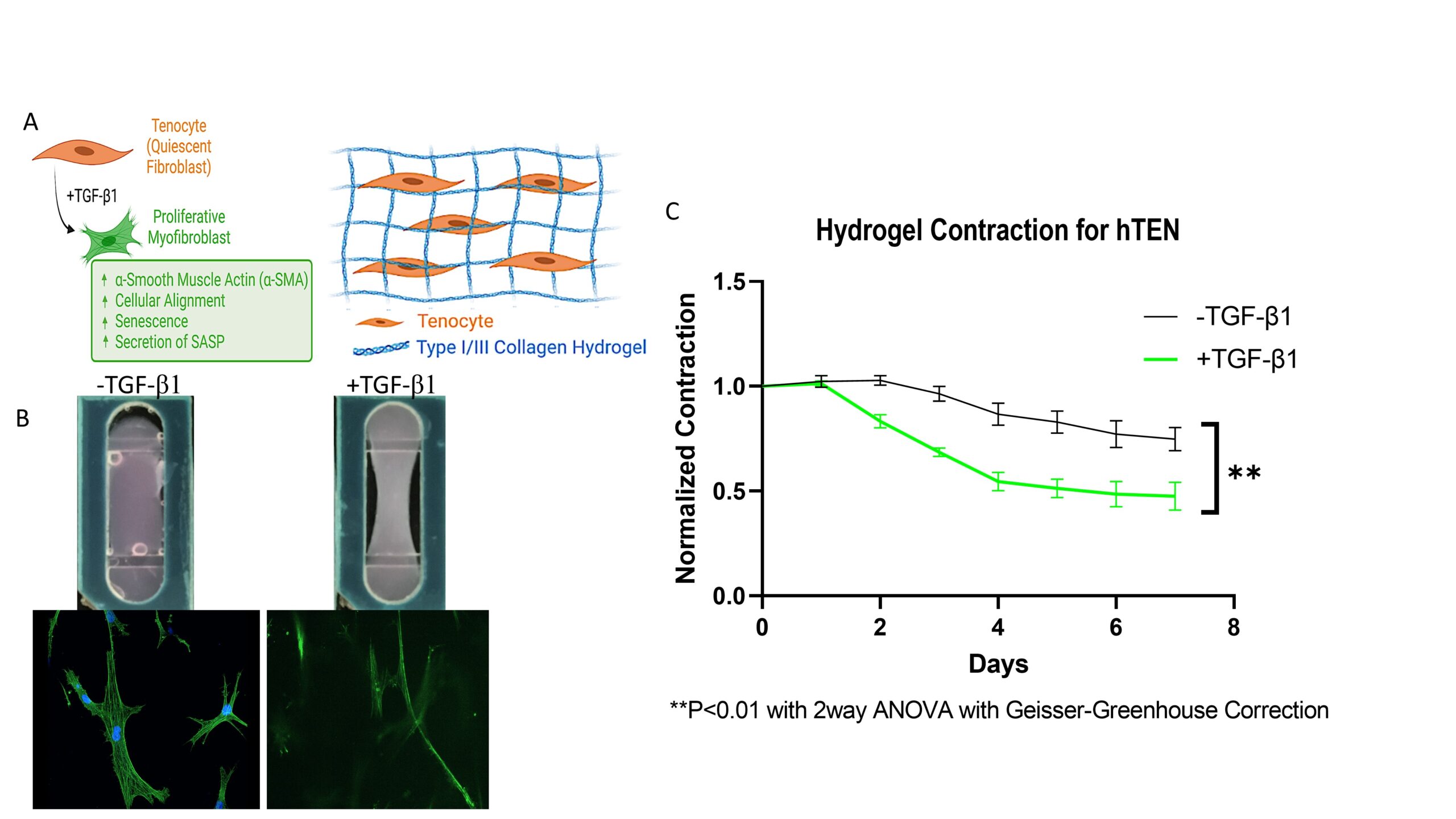
Figure 7: Myofibroblast differentiation schematic (A) and area contraction measurements (C) of tenocyte-only hydrogel. B) Representative images of area contraction in hydrogel bottom channel
Incorporation of macrophages(Mϕ) into disease model:
It is believed that macrophages are the major secretors of TGF-β1, and therefore key to the onset and progression of fibrotic disease. What is not completely understood are the distinct roles played by infiltrating macrophages into the diseased tissue and tissue-resident macrophages already in the tissue. Our platform permits to independently investigate these populations. Therefore, we sought to investigate the role of co-cultured macrophages with tenocytes, named tissue-resident macrophages in our model, and its effect on fibrotic disease initiation and progression. We started by incorporating freshly isolated CD14+ monocytes from human donors and suspended them with tenocytes at a 7:1 ratio and cultured them in MCSF+, serum-free media over 7 days in the presence and absence of TGF-β1 (see figure 6). No polarization was performed on the monocytes prior to culture to strictly identify any polarization produced through the tenocyte-monocyte signaling. Results showed high area contraction in the non-treated groups, yet low α-SMA and γh2ax expression indicating minimal myofibroblasts differentiation in the presence of unpolarized macrophages and absence of exogenous TGF-β1. This was not surprising as literature points to recruited macrophages from the vasculature as primary fibrotic and inflammatory inducers. What is notable to mention is the stark difference observed between the tenocyte only and the tenocyte/macrophage groups in their area contraction γH2AX expression, and α-SMA expression. The T/Mϕ group contracts almost 30% more than the T-only group in the absence of exogenous TGF-β1. And although the difference between the TGF-β1 in the T-only were insignificant for γh2ax expression, there was a large difference observed in the T/Mϕ group. Indicating that the co-culture of tenocytes and macrophages is significant to the induction of a senescent phenotype in myofibroblasts.
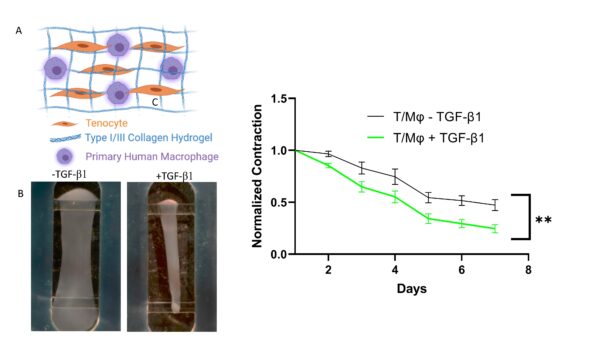
Figure 8: Tenocyte/Macrophage area contraction
Taking a closer look at the T/Mϕ group in conjunction with the T-only group we see similar contraction ratios to the TGF-β1 treated group, indicating some extent of myofibroblast activation. After staining these tissue-resident macrophages in the T/Mϕ construct we observed proinflammatory markers present in the absence of cytokine polarization or TGF-β1 treatment, indicating there is some activation occurring in the co-cultured constructs. In addition, we observed that tissue-resident macrophages can increase tissue contracture to similar extents observed with TGF-β1 treatment and they express pro-inflammatory surface markers, pointing towards an initiatory cellular phenotype.
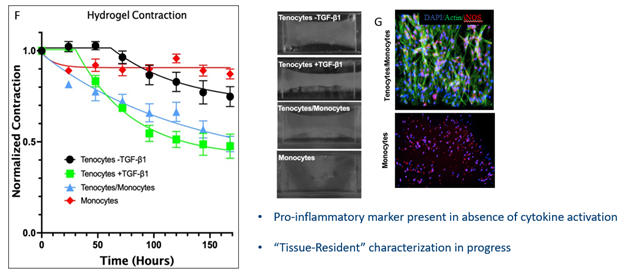
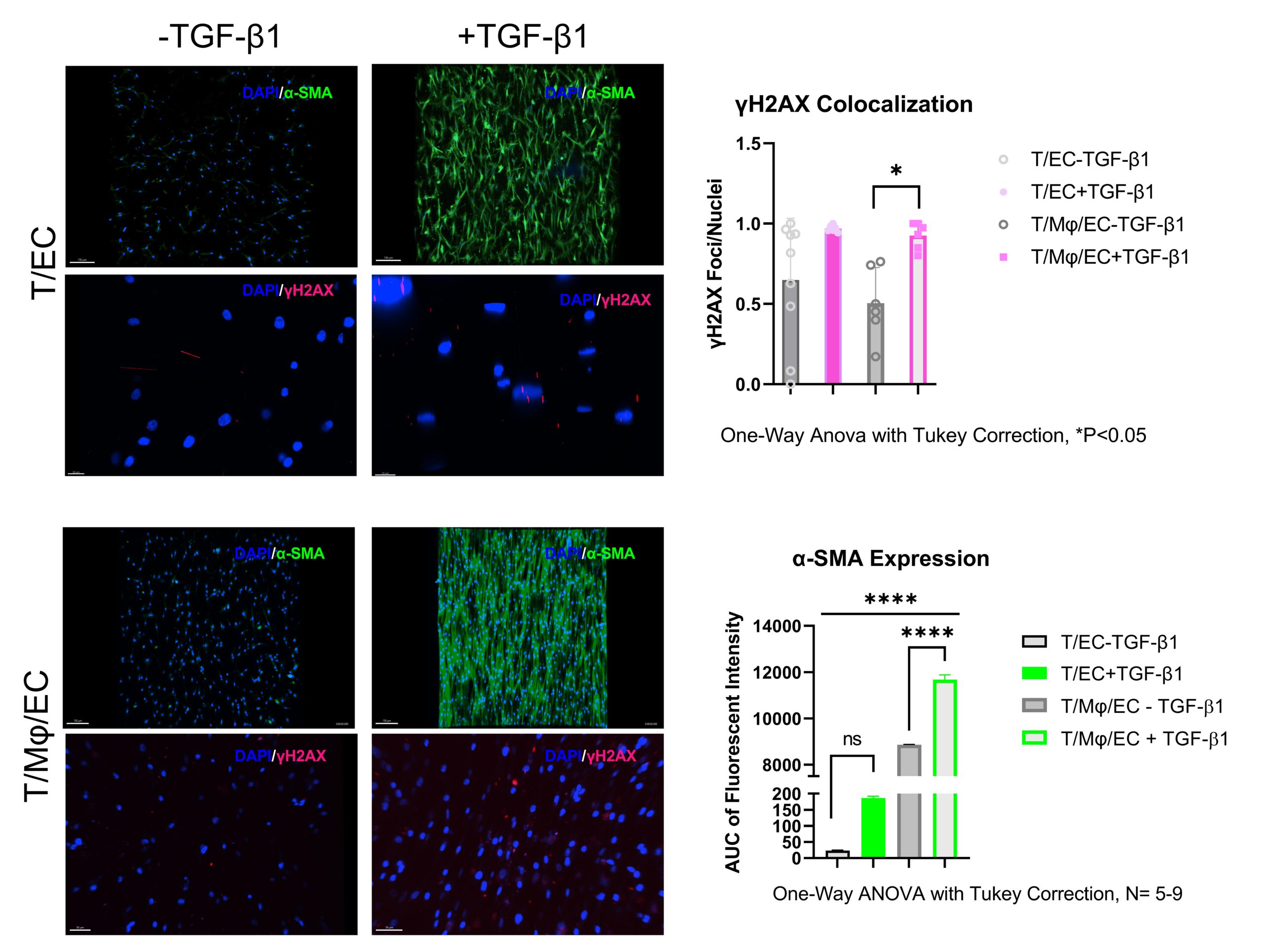
Figure 10: Alpha-SMA (green) and gh2ax (red) quantification of tenocyte-only and tenocyte/macrophage hydrogel constructs.
Live Monocyte Transmigration
As we take a closer look at the in vivo wound healing environment we see there are various other signals present including a vascular barrier which signals activated macrophages to extravasate into the tendon tissue (figure. 11). This migration propagates the inflammatory response through various cytokine signaling pathways and often leads to unresolving, chronic inflammation in tendon injuries. We incorporated this vascular barrier through our top component and introduced freshly isolated monocytes into the culture and monitored their migration into the 7-day cultured T/Mϕ hydrogel. The volumetric representation in figure 12 depicts live stained macrophages migrating through the porous barrier into the bottom hydrogel. Upon fixation, we can observe migrated monocytes are well-dispersed in hydrogel following cues from the tenocyte/macrophage-laden hydrogel, as no exogenous chemoattractants are present in the model (figure 13).
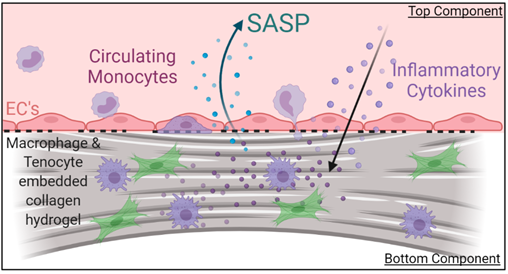
We then took the supernatant of these cultures and analyzed them against an 8-panel Luminex assay, guided from our labs data on injured mouse serum. Interestingly we found upregulation of key inflammatory cytokines only elevated in the tri-culture group from the previous slide, indicating that the present vasculature plays an important barrier and signaling role to induce transmigration and SASP secretion. Interestingly the four cytokines upregulated most in the vascularized model are highly associated with inflammatory response initiation and progression. Some of these cytokines such as Interleukin-17 blockage have proven effective strategies to treat inflammatory disorders such as systemic rheumatic diseases and serve as great treatment candidates for fibrotic diseases.
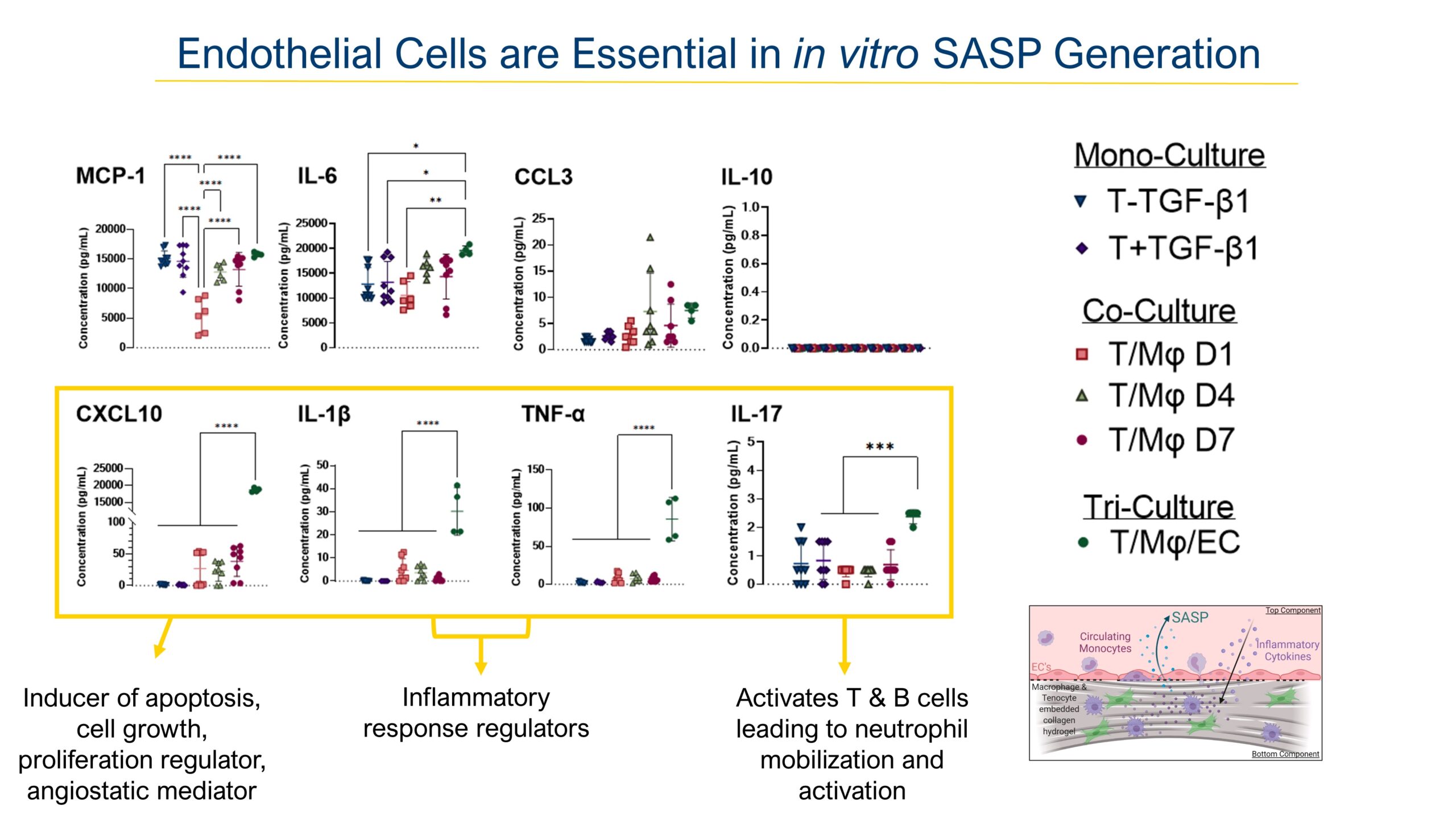
DISCUSSION:
As a summary of our findings thus far we have established a human tendon fibrotic disease model in an organ-on-a chip system complete with a vascular barrier which has shown to be vital for the representation of the fibrotic and inflammatory response in vitro. And we have also pointed to the importance of studying tissue-resident and circulating macrophages independently and in conjunction to elucidate fibrotic disease propagation. Our future effort consists of fully characterizing these macrophage populations in our system, integrating other immune cell components such as neutrophils as early responders and adapting the platform for fluid flow and mechanical actuation of the hydrogel for a more physiological response.