EC Apicobasal Polarity Short Summary: IL-8 and ICAM-1
Introduction
EC Apicobasal polarity in immunity has and will continue to be a theme of my thesis. I have presented on bits and pieces of this work over the past year or so, but now that this work is under review and more or less complete pending reviewer comments, I wanted to summarize our finds and possible future directions.
Results
Polarized Secretion of IL-8 Sequesters or Permits Neutrophil Transmigration at or to the Source of Inflammation
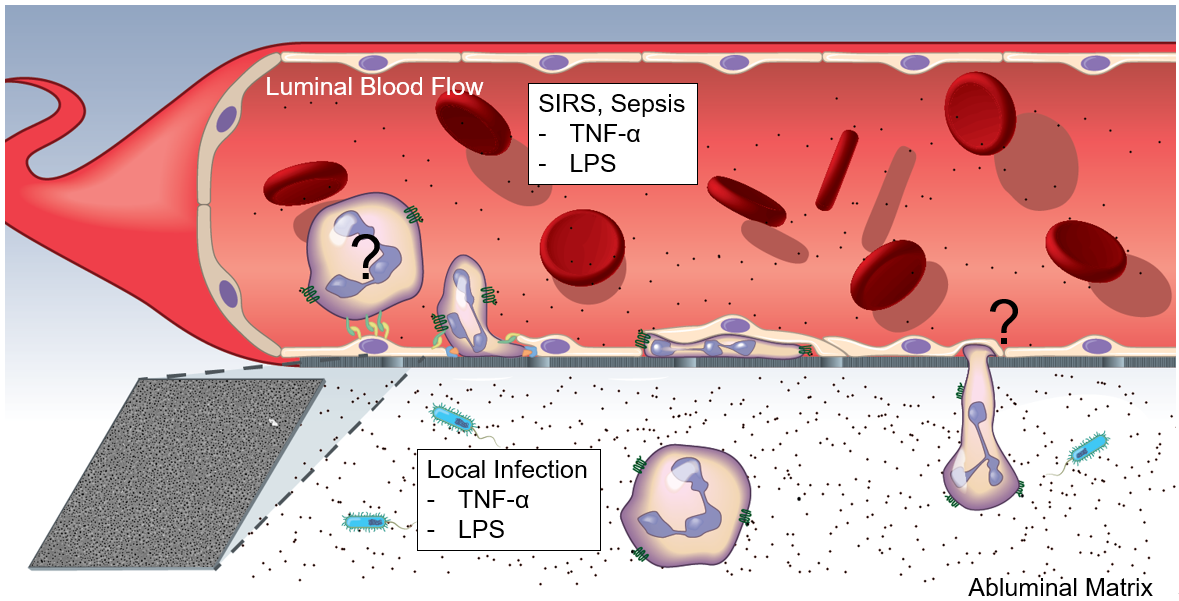
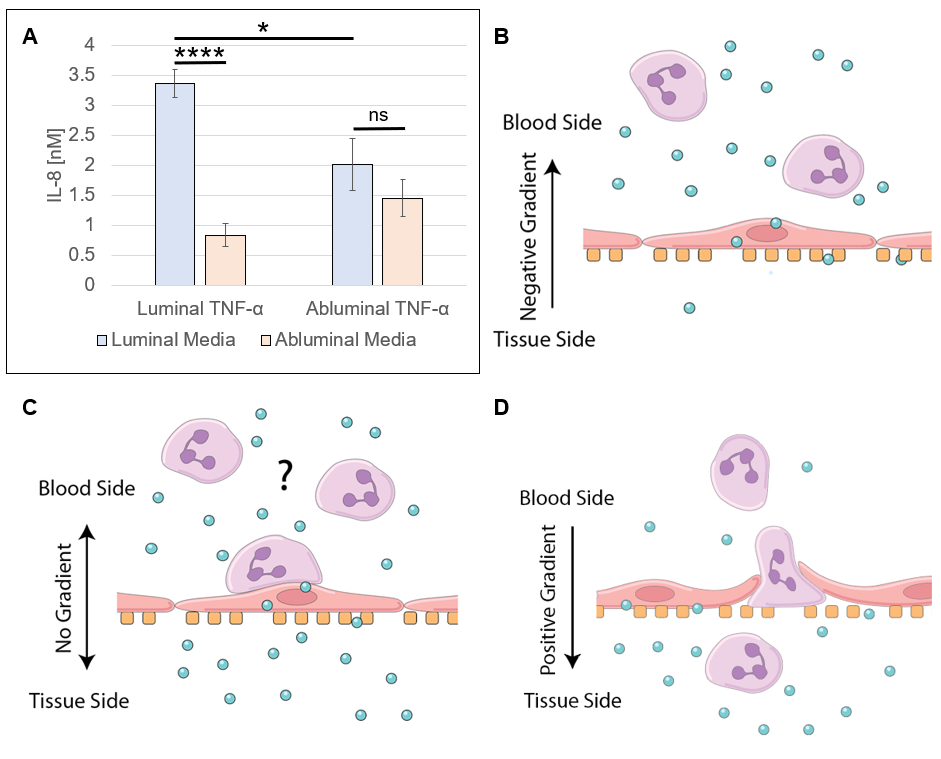
In models of systemic (luminal inflammatory cytokines) and local (abluminal inflammatory cytokines) inflammation (Figure 1), endothelial cell (EC) apicobasal polarity facilitates polarized secretion of IL-8 into the luminal and abluminal compartments. As determined by ELISA (Figure 2), systemic inflammation drives EC IL-8 secretion primarily into the luminal compartment, effectively establishing a luminally directed IL-8 gradient. Conversely, local inflammation induced a near even secretion of IL-8 into the luminal and abluminal compartments, failing to establish any trans-endothelium gradient. In a round of experiments utilizing clean media supplemented with recombinant human IL-8, its was determined that IL-8 functions to sequester neutrophils in the luminal compartment in models of systemic inflammation, and permits, BUT DOES NOT FACILITATE, neutrophil transmigration in models of local inflammation (Figure 3).
Formation of ICAM-1 Rich Membrane Ruffles in Models of Local Inflammation
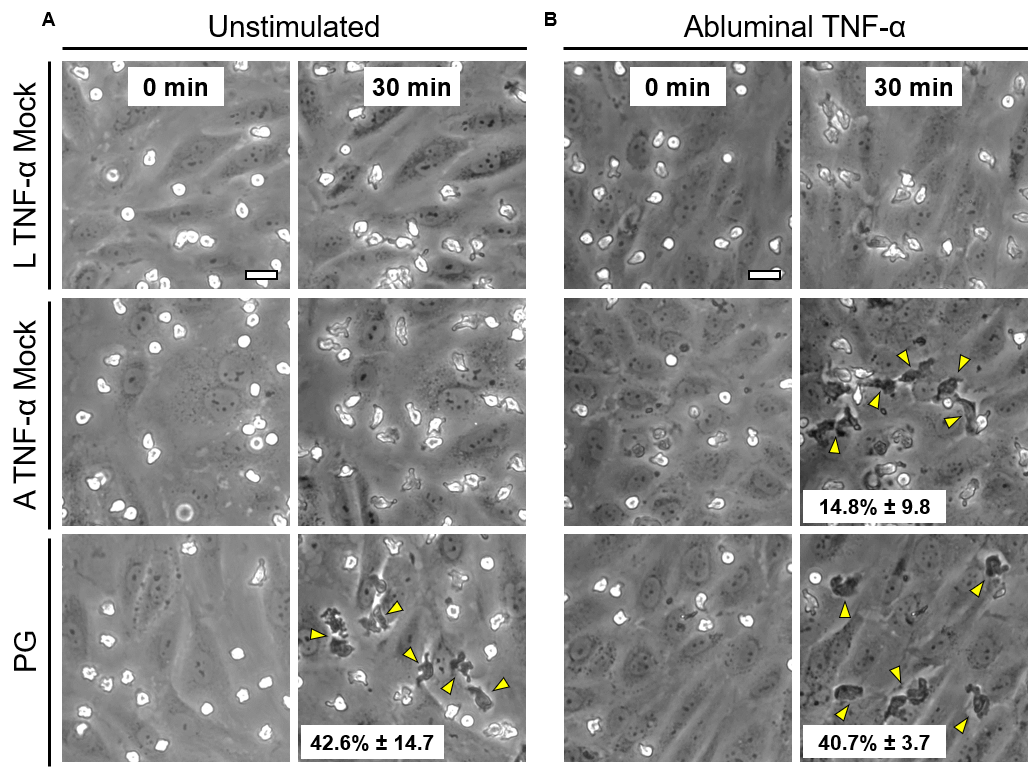
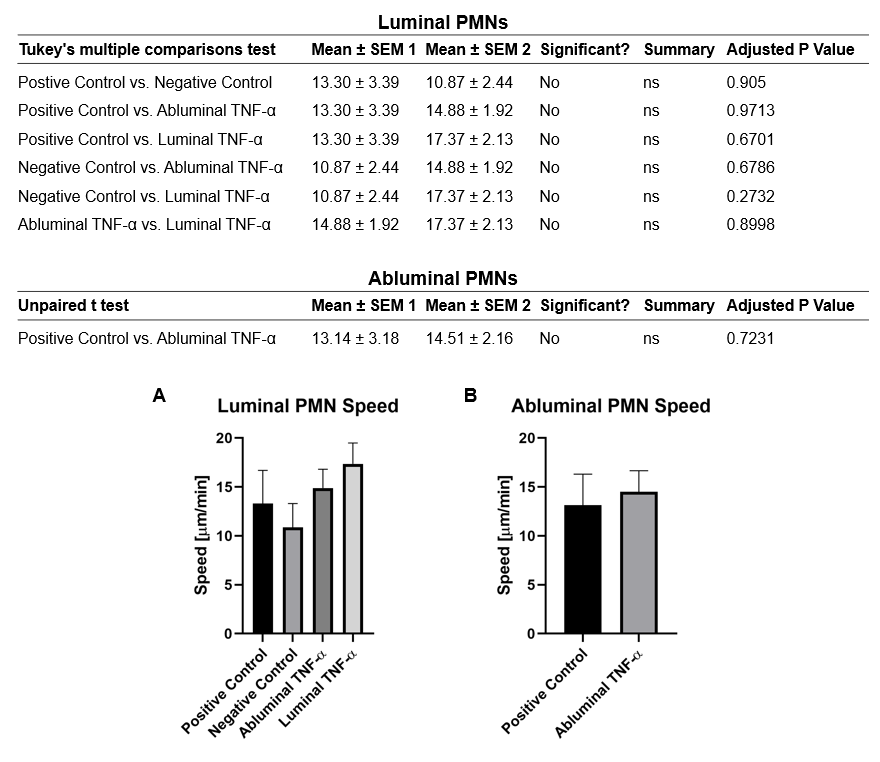
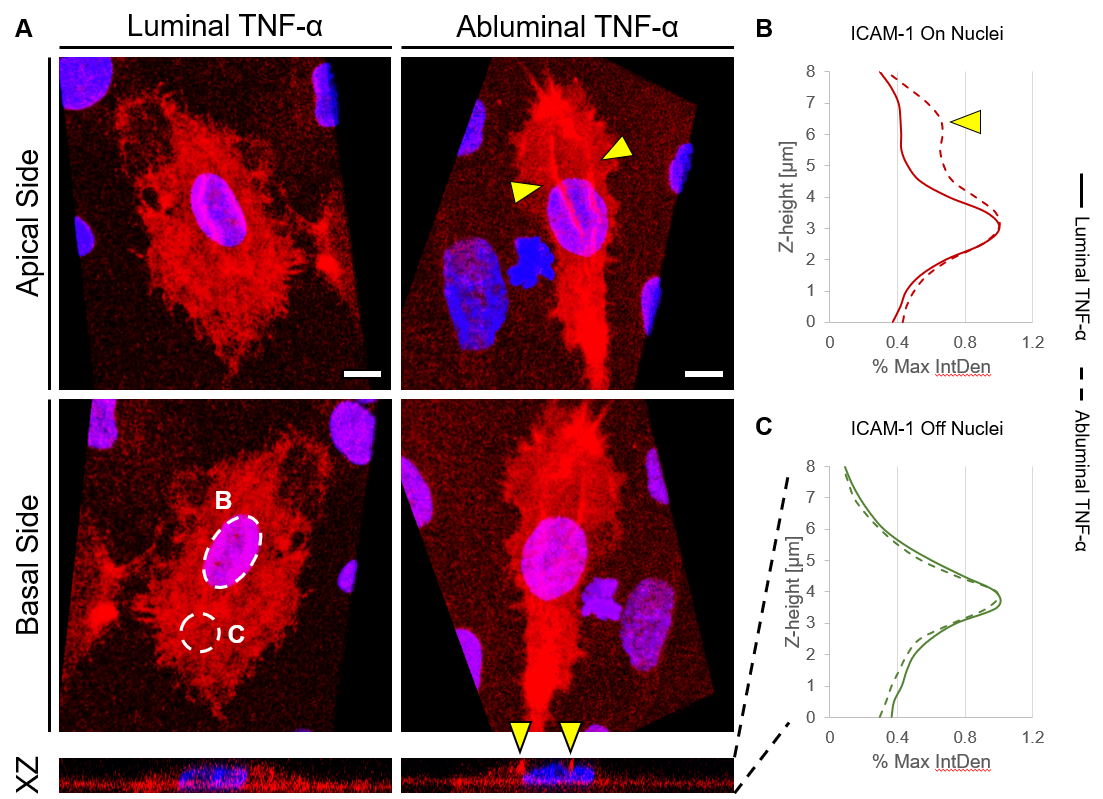
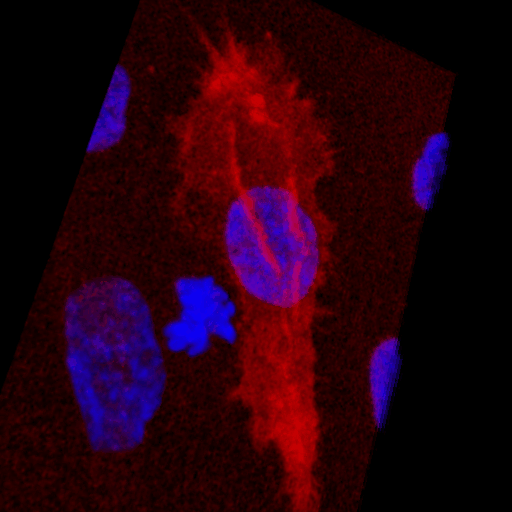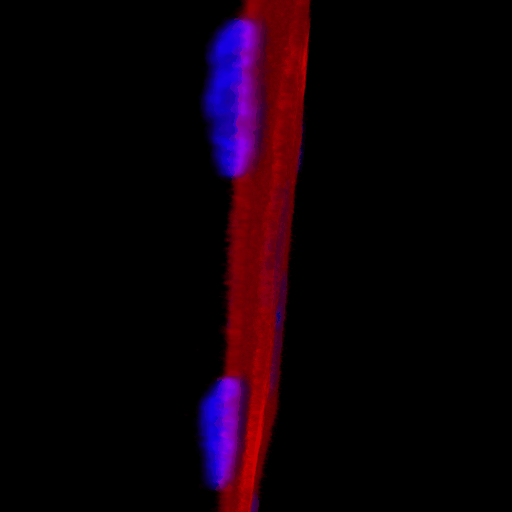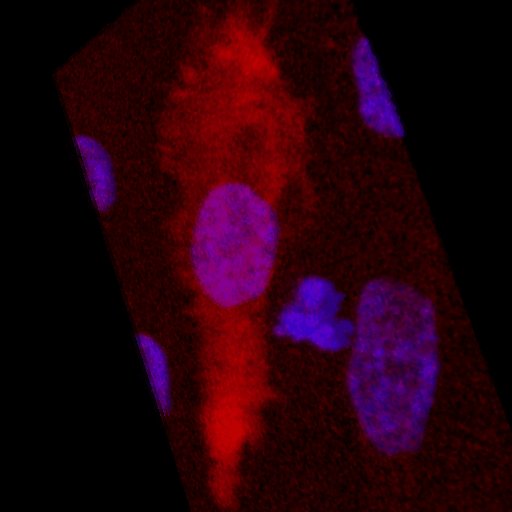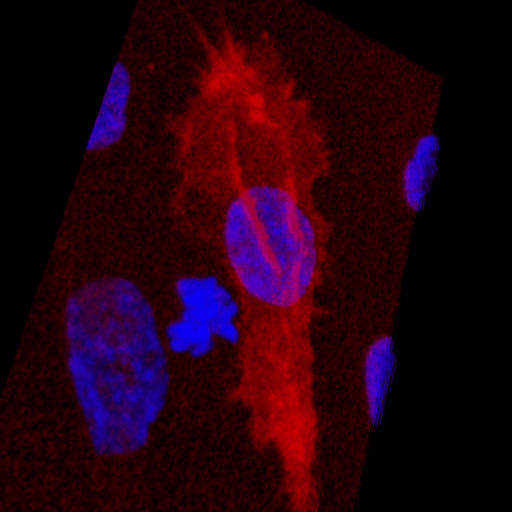
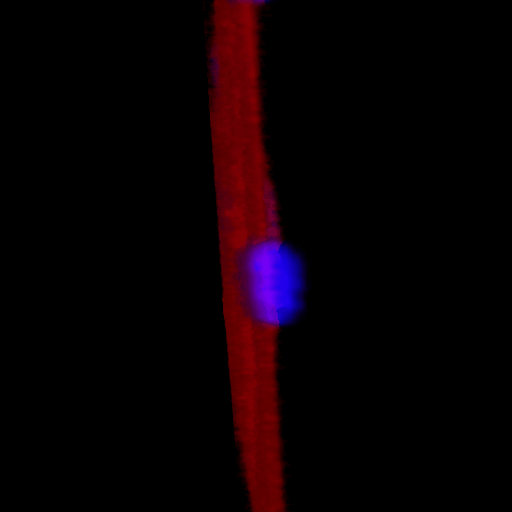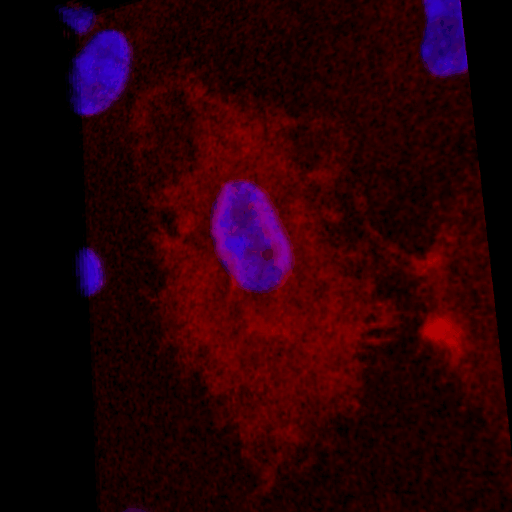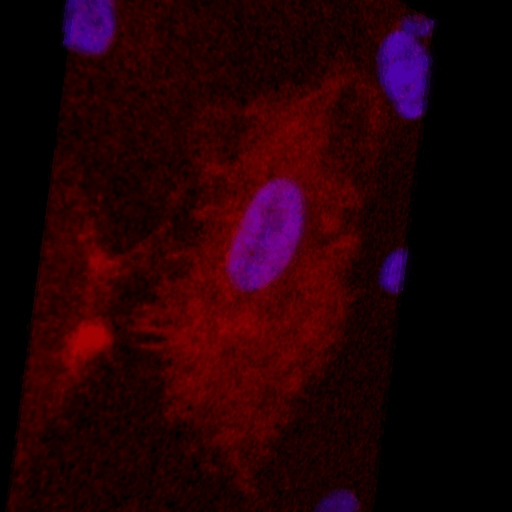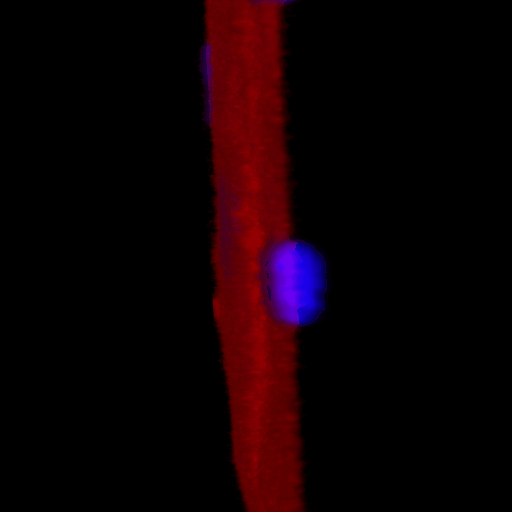
Over the course of this study, two results made us reconsider ICAM-1 for further investigation (briefly investigated via epi-fluorescence): 1) neutrophils exhibited elevated speeds in the perivascular space following EC TNF-α exposure (Figure 4), and 2) the lack of an explanation for neutrophil transmigration under the neutral IL-8 gradient produced in the model of local inflammation. The first response may be explained by an increase in ICAM-1 expression on the basal surface of abluminally stimulated ECs. Additionally, basal ICAM-1 may form a non-soluble gradient, driving neutrophil transmigration. Interestingly, while confocal analysis of EC ICAM-1 expression did reveal what appears to be basal ICAM-1 (Figure 5), local inflammation facilitated the formation of ICAM-1 rich membrane ruffles on the apical surface of ECs (Figure 6). These ruffles extended several microns above the EC surface, potentially highlighting a novel mechanism for the facilitation of neutrophil transmigration under and neutral IL-8 gradient. Ruffles appear to form in models of systemic inflammation, but do not extend into the luminal space (Figure 7).
Conclusion
EC apicobasal polarity regulates distinct immune responses to local and systemic inflammation. Future work should focus on better understanding the formation and role of ICAM-1 rich membrane ruffles.