HUVEC ICAM-1 Activation by Bacterial Extracellular Vesicles Derived From E. coli Treated with Different Antibiotics
Introduction
In prior work by Panteha Torabian and the Michel Lab (RIT’s College of Science), we have seen that treatment of E. coli with different antibiotics can result in changes in the production of bacterial extracellular vesicles (bEVs). In the context of sepsis, this is important because clinicians tend to treat patients with a broad spectrum of antibiotics. Even if the infectious bacteria are killed quickly, bacterial products such as bEVs may persist in the circulation and continue to cause proinflammatory stimulation. If certain classes of antibiotics cause the dying bacteria to produce even more bEVs, then this has the potential to exacerbate sepsis conditions. We were interested in validating whether bEVs derived from E. coli might demonstrate variable biological activity depending on the antibiotics used to treat the bacteria. Ultimately, we believe that this research could help inform clinicians about the risks of using certain antibiotics for sepsis treatment and thereby improve patient outcomes.
Methods
The following antibiotics were evaluated:
- Ciprofloxacin
- Tobramycin
- Cefepime
- Meropenem
- Amikacin
- Ceftriaxone
E. coli were grown in LB broth for 1 day prior to stimulation with the various antibiotics for 3 – 4 h. In order to standardize the treatments, twice the minimum inhibitory concentration was applied for each antibiotic. bEVs were isolated from E. coli-conditioned LB Broth via ultracentrifugation and were analyzed by nanoparticle tracking analysis. For cell stimulation experiments, HUVECs were seeded in 96-well plates at a density of 40,000 cells/cm2. The cells were treated with various concentrations of bEVs in M200 media + LVES supplement. To assess whether the effects of bEVs were mediated by lipopolysaccharide (LPS), 1 μg/ml LPS-binding protein (LBP) was spiked into the M200 + LVES during the treatment. After 16 – 18 h, HUVEC inflammatory activation was assessed by live-labeling the cells with an anti-ICAM-1 antibody for 15 min prior to fixation with 4% formaldehyde and subsequent incubation with a fluorescent secondary antibody. The ICAM-1 fluorescent signal obtained from microscopy images was averaged and normalized to the number of counterstained nuclei detected in each image.
Results and Discussion
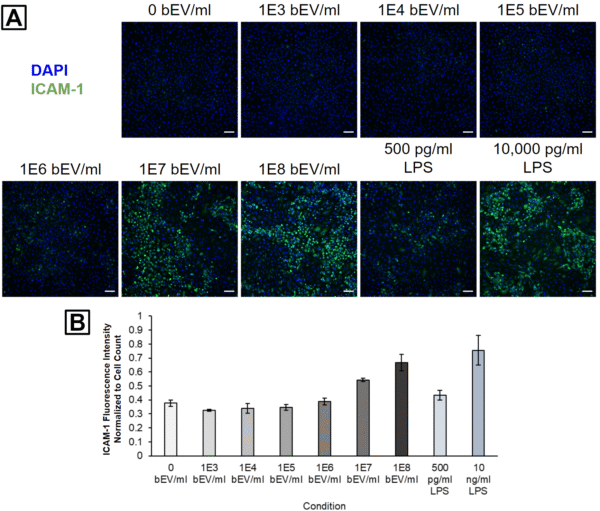
We first examined a wide dosing range with bEVs obtained from healthy E. coli (no antibiotic treatment). HUVEC ICAM-1 activation appeared elevated after treatment with 1E6 – 1E8 bEVs/ml, and these activation levels were comparable to those obtained with 500 or 10,000 pg/ml LPS (Figure 1).
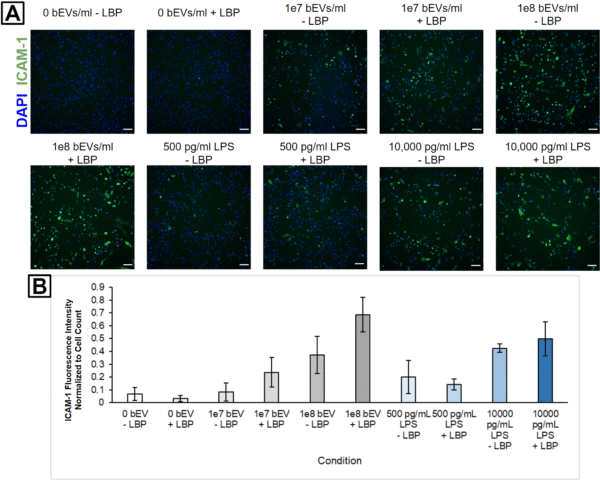
We were curious if the primary proinflammatory mediator was LPS associated with the bEVs. To investigate this, we added 1 μg/ml LBP to the HUVEC culture media since LBP forms a complex with LPS that makes it more detectable by toll-like receptors on the cell surface. It is worth noting, however, that our media likely already has a baseline level of LBP due to the LVES supplement. The presence of additional LBP led to increased ICAM-1 activation when HUVECs were treated with 1E7 and 1E8 bEVs/ml, but did not seem to increase activation when the cells were treated with 500 or 10,000 pg/ml LPS (Figure 2). We believe this indicates that bEV-associated LPS may play a role in HUVEC stimulation, but that the low amount of LBP in the media may be a limiting reagent. Conversely, when we treated cells with pure LPS, it seems like the LPS itself behaves like a limiting reagent since the addition of more LBP did not change the cell response. This could mean that the bEVs possess much higher concentrations of LPS or that their LPS is oriented in a manner that increases its ability to interact with recipient cells. We plan to re-investigate this behavior in the future using LVES-free media to remove the confounding effects of the baseline level of LBP.
We treated the E. coli with the 6 antibiotics and isolated bEVs from the conditioned LB broth. Nanoparticle tracking analysis demonstrated differences in the recovered concentrations and mean sizes of the bEVs (Table 1). Notably, Ceftriaxone demonstrated a 5.2-fold increase over the control (no antibiotics) while Amikacin produced a 3.7-fold increase over the control.
Table 1. Nanoparticle tracking analysis data of bEVs prepared from E. coli treated with different antibiotics. Concentrations were recorded from 1:1,000 dilutions and back-calculated. Data represents the average from three 30 s videos +/- SEM.
| Condition | Concentration (particles/ml) | Mean Particle Size (nm) |
| Control (no antibiotics) | 1.77e11 +/- 1.80e10 | 165.0 +/- 3.2 |
| Amikacin | 6.55e11 +/- 3.31e10 | 170.6 +/- 1.3 |
| Cefepime | 4.74e11 +/- 1.73e10 | 179.4 +/- 2.6 |
| Ceftriaxone | 9.33e11 +/- 7.95e09 | 158.1 +/- 5.2 |
| Ciprofloxacin | 2.53e11 +/- 4.37e09 | 182.2 +/- 2.8 |
| Meropenem | 4.61e11 +/- 2.19e10 | 176.2 +/- 1.4 |
| Tobramycin | 2.00e11 +/- 6.45e09 | 189.5 +/- 3.3 |
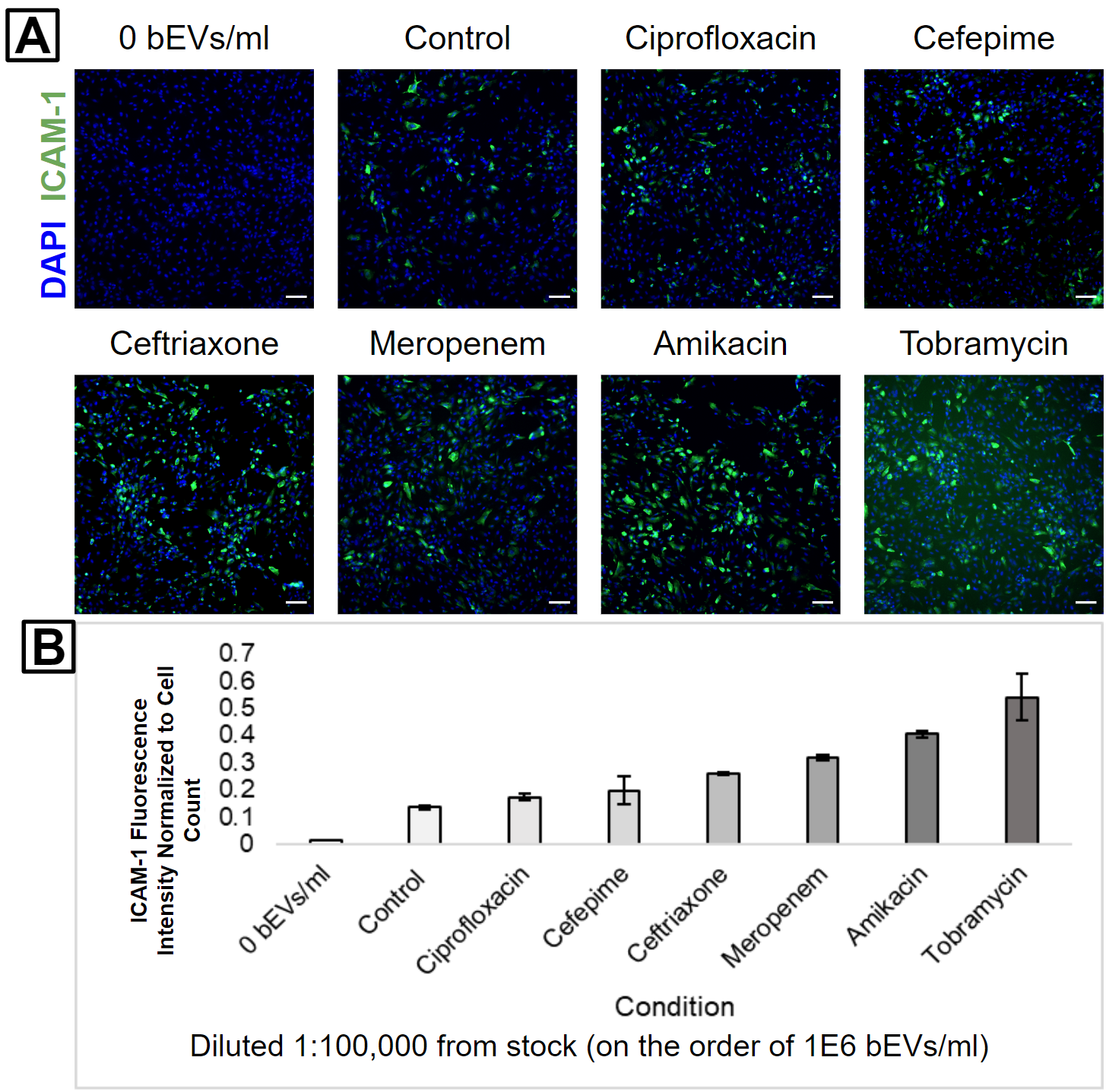
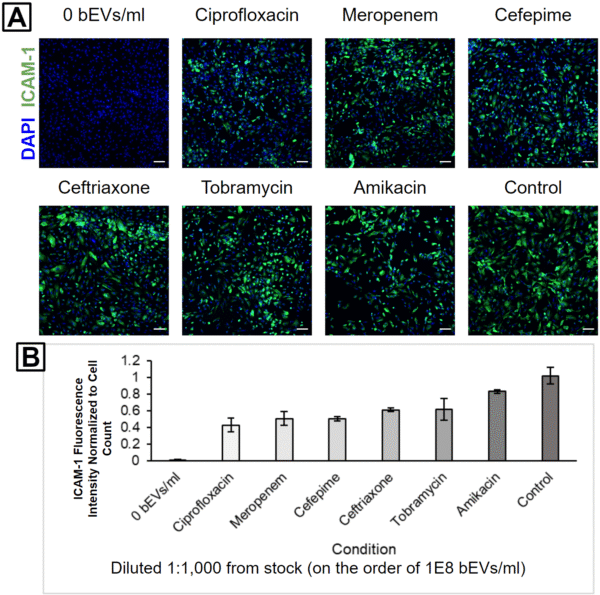
HUVECs were subsequently treated with 1:1,000 (Figure 3), 1:10,000 (Figure 4), or 1:100,000 (Figure 5) dilutions from each bEV stock. This was performed in order to keep the relative concentrations the same between the different bEV preparations. Interestingly, the condition causing the highest ICAM-1 activation changed with each dilution level. The control bEVs caused the highest ICAM-1 activation for the 1:1,000 dilution, Amikacin bEVs caused the highest ICAM-1 activation for the 1:10,000 dilution, and Tobramycin bEVs caused the highest ICAM-1 activation for the 1:100,000 dilution.
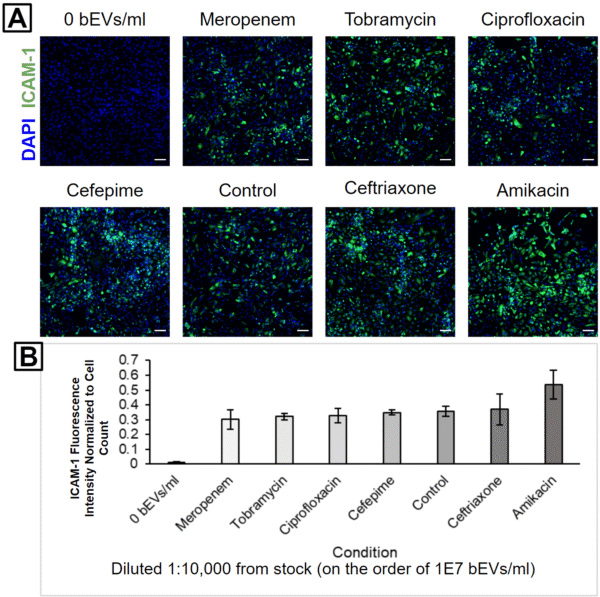
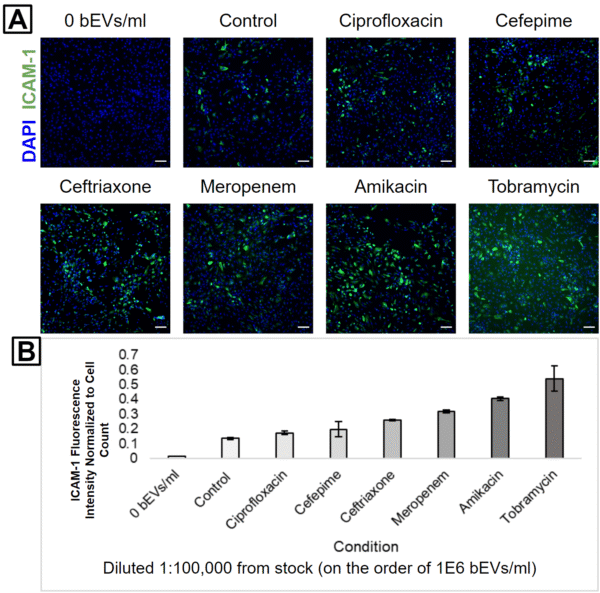
It is important to note that despite these apparent differences, the actual ICAM-1 fluorescence values were clustered pretty tightly together for each dilution level. Almost every antibiotic bEV was internally consistent; their capacity to activate ICAM-1 always decreased as the bEV concentrations were reduced (Figure 6). The only big exception was Tobramycin, but the high background signal in its 1:100,000 fluorescence image suggests that this may be due to labeling issues (Figure 5A). Furthermore, the experiment was unreplicated, and some of the “order-swapping” trends might be erased by the inclusion of more technical replicates.

One interesting finding was that Amikacin bEVs were always one of the top 2 ICAM-1 activators. This is notable since there were fewer bEVs in the Amikacin-treated sample compared to the Ceftriaxone-treated sample. In other words, the Amikacin bEVs would have to possess higher biological activity than the Ceftriaxone bEVs in order to produce a higher HUVEC ICAM-1 activation. This is our first evidence that different antibiotic treatments can modulate both the concentration and biological activity of bEVs from E. coli.